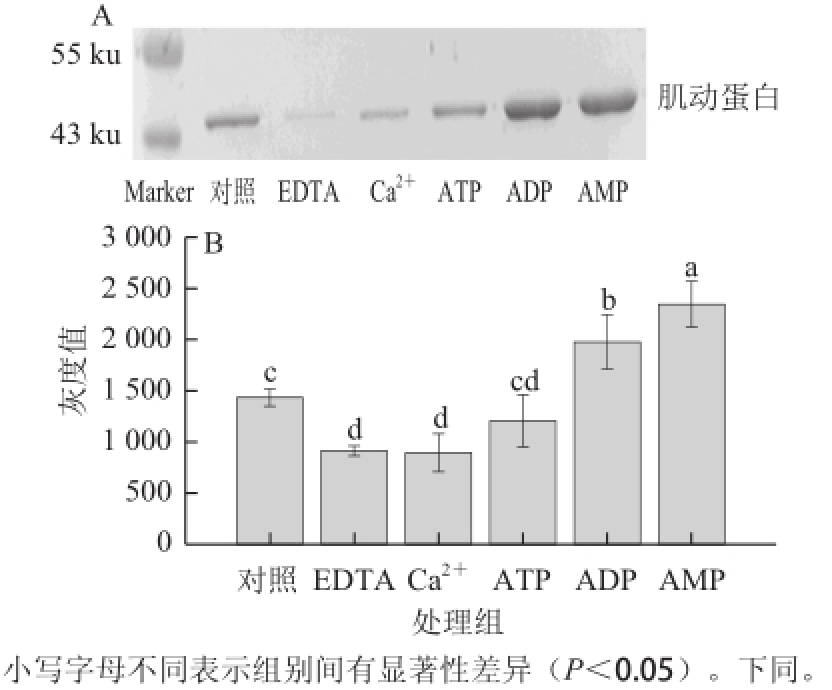
图1 EDTA、Ca2+、ATP、ADP、AMP对肌动球蛋白解离影响的Western blotting转印图(A)和定量分析图(B)
Fig.1 Western blotting (A) and quantitative analysis (B) of actomyosin dissociation as affected by EDTA, Ca
2+, ATP, ADP and AMP
邓少颖 1,2,王道营 1,*,张牧焓 1,卞 欢 1,吴海虹 1,诸永志 1,耿志明 1,刘 芳 1,徐为民 1
(1.江苏省农业科学院农产品加工研究所,江苏 南京 210014;2.南京农业大学,教育部肉品加工与质量控制重点实验室,江苏 南京 210095)
摘 要:为了解促进肌动球蛋白解离的因素,从鸭胸肉中提取肌动球蛋白,研究Ca 2+、三磷酸腺苷(adenosine triphosphate,ATP)及其降解产物对肌动球蛋白的解离效果。通过蛋白质免疫印迹技术测定肌动蛋白含量的变化来研究肌动球蛋白的解离情况。研究发现,7.5~64 mmol/L Ca 2+对肌动球蛋白的解离无促进作用(P>0.05),而Ca 2+浓度升高到200 mmol/L时,能显著促进肌动球蛋白的解离(P<0.05);单独的ATP对肌动球蛋白的解离无促进作用(P>0.05);Ca 2+和ATP共同作用于肌动球蛋白后,可显著促进肌动球蛋白的解离(P<0.05);二磷酸腺苷(adenosine diphosphate,ADP)、一磷酸腺苷(adenosine monophosphate,AMP)均可显著促进肌动球蛋白的解离(P<0.05),且不同浓度处理组间无显著性差异(P>0.05)。因此,可以推断ADP、AMP以及Ca 2+和ATP的共同作用对肌动球蛋白的解离有促进作用。
关键词:鸭肉;肌动球蛋白;蛋白质免疫印迹;解离;肌动蛋白
肉中含有丰富的蛋白质和维生素,是人机体获取氨基酸的主要来源,肉中还含有较多的含氮浸出物,赋予肉品较好的风味。在选购肉品时,嫩度已成为一个重要的评定指标 [1-3],研究肉的成熟机制及改善肉的嫩度已成为肉品领域研究的热点问题。
肌原纤维由粗丝和细丝组成,粗丝主要由肌球蛋白组成,又称为“肌球蛋白丝”,而细丝主要是由肌动蛋白分子组成,并辅以结合原肌球蛋白和肌钙蛋白,因此又称“肌动蛋白丝”,肌球蛋白和肌动蛋白约占肌原纤维的70%~75% [4]。肌原纤维是肌肉的伸缩装置,当肌肉收缩时,肌动蛋白和肌球蛋白结合形成肌动球蛋白横桥,肌节变短;松弛时,肌动球蛋白横桥断裂,肌节变长。Takahashi等 [5]研究指出:宰后成熟过程中肌节长度的增加是由于肌动蛋白和肌球蛋白结合状态以及结合程度的改变。Koohmaraie [6]推测宰后成熟过程中肌球蛋白和肌动蛋白之间相互作用的弱化可能对于肉品嫩度的改善起到了重要作用。Okitani等 [7]研究加热过程中肉嫩度的变化规律时发现,当肉嫩度相对较好时,有大量的肌动球蛋白发生解离。有研究者认为,成熟过程肌浆中高浓度的Ca 2+激活钙蛋白酶后作用于肌原纤维骨架蛋白,使之发生降解,直接或间接的影响肌动球蛋白横桥的连接 [8-10]。在宰后成熟过程中,细胞无氧酵解产生的三磷酸腺苷(adenosine triphosphate,ATP)不足以补充生化反应的消耗,使得细胞内的ATP急剧降低,但其水解产物二磷酸腺苷(adenosine diphosphate,ADP)、一磷酸腺苷(adenosine monophosphate,AMP)却不断积累 [11]。肌球蛋白和肌动蛋白的结合状态及结合程度可能受到ATP及其降解产物的影响。Okitani等 [12]研究发现从鸡、牛、猪骨骼中提取的肌动球蛋白溶液,经AMP孵育后,肌动球蛋白发生了解离。
因此,本实验拟对Ca 2+、ATP以及其降解产物能否引起鸭肉中肌动球蛋白的解离进行研究,并为肌动球蛋白横桥的生成和断裂与肉品嫩度之间的关系提供理论基础研究。
1.1 材料与试剂
肉用麻鸭,购自南京孝陵卫农贸市场,随机选取生长60 d左右、大小相近的健康肉用麻鸭。
ATP、ADP、AMP、抗兔骨骼肌肌动蛋白多克隆抗体、羊抗兔IgG、三羟甲基氨基甲烷(Tris)(超纯级)美国Sigma公司;DAB辣根过氧化物酶显色试剂盒、CaCl 2(分析纯) 南京巴傲得生物科技公司;预染宽范围标准蛋白(分子质量大小范围14.4~116 ku) 加拿大Fermentas公司;丙烯酰胺、甲叉双丙烯酰胺(分析纯) 丁贝生物科技有限公司。
1.2 仪器与设备
M124A电子分析天平 意大利BEL公司;SPX-250B-Z生化培养箱 上海博讯有限公司;Mini-PROTEAN ☒Tetra Cell垂直电泳系统、Semi-Dry转印仪 美国Bio-Rad公司;T-25数显匀浆机 德国IKA公司;JS-680C全自动凝胶成像分析仪 上海培清科技有限公司。
1.3 方法
1.3.1 肉样处理
将选取的麻鸭宰杀前12 h禁食、前3 h禁水后,采取颈部左侧割断其动脉静脉放血的方式实行宰杀(宰杀时未采取任何击昏方式),放血完成后立即取出胸大肌,置于4 ℃冰箱中成熟12 h后,去除肌膜、结缔组织、脂肪后切碎成肉糜状,待用。
1.3.2 肌动球蛋白的提取方法
参考Benjakul等 [13]的方法从鸭胸肉中提取肌动球蛋白,并稍做修改。将处理后的2 g肉糜样品置于20 mL的Weber-Edsall提取液(0.6 mol/L KCl,0.01 mol/L Na 2CO 3,0.04 mol/L NaHCO 3,pH 7.2)中,于冰浴中15 000 r/min匀浆3 次(每次30 s,间隔30 s)。将匀浆液置于4 ℃恒温摇床培养箱中提取24 h后,加入40 mL蒸馏水稀释提取液中KCl的浓度,使其终浓度为0.2 mol/L,4 ℃摇床振荡60 min后将溶液进行离心分离,离心条件为:4 ℃、15 000×g,20 min;弃去上清液后,重新加入20 mL Weber-Edsall提取液,再加入40 mL蒸馏水稀释,用两层尼龙网过滤弃去不溶物质后,4 ℃摇床振荡60 min后再进行离心分离(4 ℃、15 000×g,20 min),弃去上清液,将沉淀溶解于5 mL KCl-Tris溶液中(0.6 mol/L KCl、20 mmol/L Tris-HCl,pH 7.2),考马斯亮蓝法测定蛋白质量浓度后,调整蛋白终质量浓度为6 mg/mL,随后立即对肌动球蛋白溶液进行相应的实验处理。
1.3.3 溶液的配制
ATP溶液:20 mmol/L Tris-HCl(pH 7.2),x mmol/L ATP(x为ATP浓度,x=8、16、24、32)。
ADP、AMP溶液:同ATP溶液,分别以ADP、AMP替换ATP。
CaCl 2溶液:20 mmol/L Tris-HCl(pH 7.2),x mmol/L CaCl 2(x为CaCl 2浓度,x=7.5、16、24、32、64、200)。
乙二胺四乙酸(ethylenediaminetetraacetic acid,EDTA)溶液:20 mmol/L Tris-HCl(pH 7.2),16 mmol/L EDTA。
1.3.4 实验设计
以提取的肌动球蛋白为实验原料,验证Ca 2+、EDTA、ATP、ADP、AMP等对其解离的影响。共设6 个处理组。每组按以下要求进行处理,设置3 个重复。
处理1:分别准确吸取1 mL的肌动球蛋白溶液,放入10 mL离心管中,分别加入2 mL的20 mmol/L Tris-HCl(pH 7.2)(对照组)、16 mmol/L的EDTA、Ca 2+、ATP、ADP、AMP。
处理2:不同浓度的Ca 2+对肌动球蛋白的解离作用。分别准确吸取1 mL的肌动球蛋白溶液,放入10 mL离心管中,对照组加入2 mL 20 mmol/L Tris-HCl(pH 7.2),处理组依次加入2 mL 7.5、16、24、32、64、200 mmol/L的Ca 2+。
处理3:不同浓度的ATP对肌动球蛋白的解离作用。分别准确吸取1 mL的肌动球蛋白溶液,放入10 mL离心管中,对照组加入2 mL 20 mmol/L Tris-HCl(pH 7.2),处理组依次加入2 mL 8、16、24、32 mmol/L的ATP。
处理4:不同浓度的ADP对肌动球蛋白的解离作用。分别准确吸取1 mL的肌动球蛋白溶液,放入10 mL离心管中,对照组加入2 mL 20 mmol/L Tris-HCl(pH 7.2),处理组依次加入2 mL 8、16、24、32 mmol/L的ADP。
处理5:不同浓度的AMP对肌动球蛋白的解离作用。分别准确吸取1 mL的肌动球蛋白溶液,放入10 mL离心管中,对照组加入2 mL 20 mmol/L Tris-HCl(pH 7.2),处理组依次加入2 mL 8、16、24、32 mmol/L的AMP。
处理6:Ca 2+与ATP、ADP或AMP共同作用对肌动球蛋白解离的影响。分别准确吸取1 mL的肌动球蛋白溶液,放入10 mL离心管中,于各试管中先加入1 mL 16 mmol/L Ca 2+后,对照组加入1 mL 20 mmol/L Tris-HCl(pH 7.2),3 个处理组分别加入1 mL 16 mmol/L ATP、ADP或AMP。
上述6 个处理组加试剂充分混匀后,置于4 ℃恒温摇床培养箱中反应24 h后,于4 ℃条件下进行离心(15 000×g,20 min)分离,取上清液,制备电泳样品。
1.3.5 肌动球蛋白解离的检测方法
将含有肌动蛋白和肌动球蛋白的中性KCl溶液进行离心分离,肌动蛋白存在于上清液中,肌动球蛋白存在于沉淀中。通过Western blotting测定上清液中肌动蛋白含量的变化来研究肌动球蛋白的解离情况 [14]。采用12%的分离胶、5%的浓缩胶进行蛋白电泳(m(丙烯酰胺):m(甲叉双丙烯酰胺)=36.5:1),每孔等体积上样20 ☒L,200 V恒压电泳约70 min。电泳结束后,切割分离胶中所需蛋白条带并在转膜液中浸泡约15 min后,转印到聚偏二氟乙烯(polyvinylidene fluoride,PVDF)膜上。转印后的PVDF膜在TBST(Tris-buffered-saline with Tween)溶液(50 mmol/L Tris-HCl、150 mmol/L NaCl、5 mmol/L KCl、体积分数为0.05% Tween-20,pH 7.4)中漂洗5 次,每次5 min。然后将带有蛋白的PVDF膜在含有质量分数5%脱脂奶粉的TBST溶液中室温孵育2 h。经TBST溶液漂洗3 次后,再将PVDF膜与一抗(抗兔骨骼肌肌动蛋白多克隆抗体,TBST溶液1:1 000稀释)结合,4 ℃孵育12 h,然后膜用TBST溶液再次漂洗3 次后与二抗(辣根过氧化酶连接的羊抗兔IgG,TBST溶液1:5 000稀释)结合,室温孵育2 h,TBST溶液漂洗3 次后,用DAB显色剂对膜进行显色处理,并使用凝胶成像仪拍照。Quantity One软件扫描蛋白免疫印迹条带,并进行数据分析。
1.4 数据分析
凝胶成像仪拍照后的图片(3个平行),Quantity One软件扫描蛋白免疫印迹条带所得的数据,用SPSS18.0进行One-way ANOVA分析,用Duncan’s multiple range tests模型进行显著性分析,显著水平为P<0.05。
图1 EDTA、Ca2+、ATP、ADP、AMP对肌动球蛋白解离影响的Western blotting转印图(A)和定量分析图(B)
Fig.1 Western blotting (A) and quantitative analysis (B) of actomyosin dissociation as affected by EDTA, Ca
2+, ATP, ADP and AMP
2.1 EDTA、Ca 2+、ATP、ADP、AMP对肌动球蛋白解离的影响由图1A可知,EDTA、Ca 2+处理组的肌动蛋白含量较对照组少,ATP处理组与对照组相差不大,而ADP、AMP处理组中的肌动蛋白含量较对照组增加。由图1B可知,EDTA、Ca 2+处理组的肌动蛋白条带灰度较对照组显著降低(P<0.05);ATP处理组的肌动蛋白条带灰度与对照组无显著性差异(P>0.05);ADP、AMP处理组均较对照组显著性增加(P<0.05),经AMP处理后肌动球蛋白解离的最多,表明AMP对肌动球蛋白的解离效果最好。
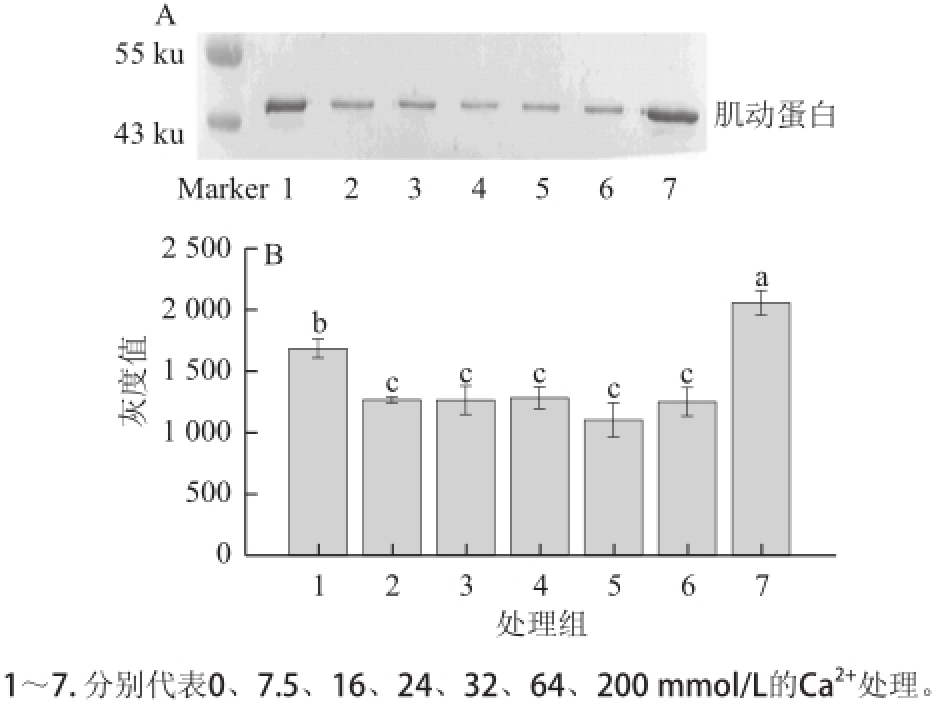
图2 Ca 2 Ca
2+2+对肌动球蛋白解离影响的Western blotting转印图(A)和定
A量分析图(B)B
Fig.2 Western blotting (A) and quantitative analysis (B) for the effects of Ca
2+on actomyosin dissociation
2.2 不同浓度的Ca 2+对肌动球蛋白解离的影响由图2A可知,除200 mmol/L Ca 2+处理组外,其他处理组中肌动蛋白含量均较对照组减少。由图2B可知,
7.5 、16、24、32、64 mmol/L Ca 2+处理组中肌动蛋白条带灰度较对照组显著降低(P<0.05);200 mmol/L Ca 2+处理组中的肌动蛋白条带灰度较对照组及其他处理组显著增加(P<0.05)。
2.3 不同浓度的ATP对肌动球蛋白解离的影响
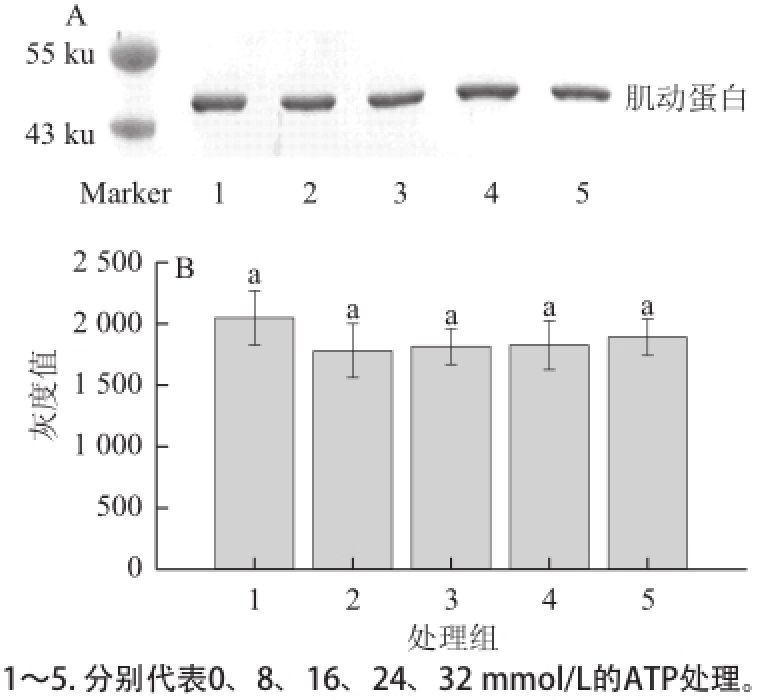
图3 ATP对肌动球蛋白解离影响的Western blotting转印图(A)和定量分析图(B)B
Fig.3 Western blotting (A) and quantitative analysis (B) for the effect of ATP on actomyosin dissociation
由图3A可知,经不同浓度的ATP处理后,肌动蛋白含量未发生明显的变化。由图3B可知,各个处理组与对照组肌动蛋白灰度均无显著性差异(P>0.05)。
2.4 不同浓度的ADP对肌动球蛋白解离的影响
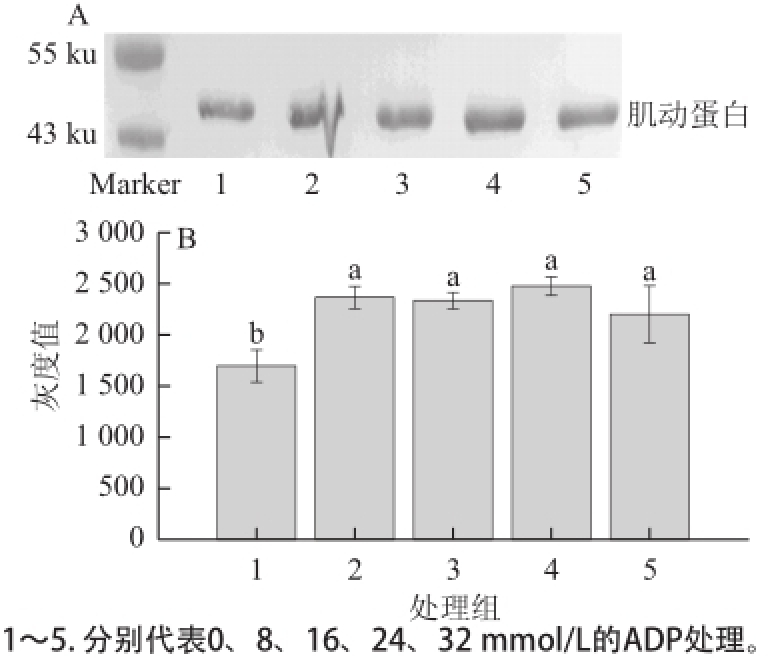
图4 ADP对肌动球蛋白解离影响的Western blotting转印图(A)和定量分析图(B)B
Fig.4 Western blotting (A) and quantitative analysis (B) for the effect of ADP on actomyosin dissociation
由图4A可知,各个处理组的肌动蛋白含量较对照组略有增加。由图4B可知,各个处理的肌动蛋白条带灰度均较对照组显著增加(P<0.05);但各个处理组间则无显著差异(P>0.05)。
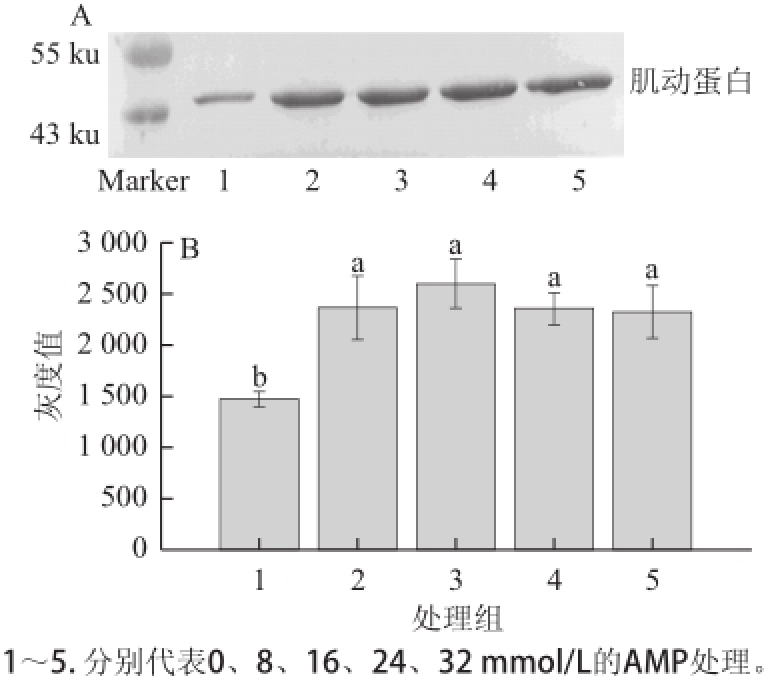
图5 AMP对肌动球蛋白解离影响的Western blotting转印图(A)和定量分析图(B)B
Fig.5 Western blotting (A) and quantitative analysis (B) for the effects of AMP on actomyosin dissociation
2.5 不同浓度的AMP对肌动球蛋白解离的影响由图5A可知,各个处理组的肌动蛋白含量均明显高于对照组。由图5B可知,经不同浓度的AMP处理后,肌动蛋白条带灰度均较对照组显著增加(P<0.05);各个处理组间则无显著性差异(P>0.05)。
2.6 Ca 2+与ATP、ADP或AMP共同作用对肌动球蛋白解离的影响
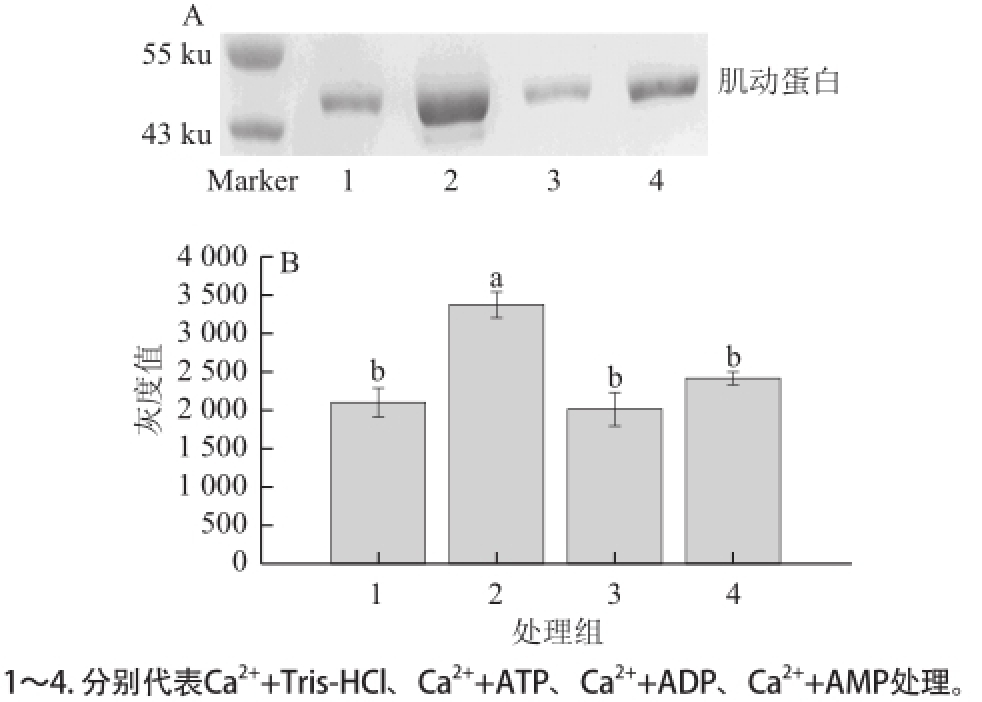
图 6 Ca
2+和ATP、ADP、AMP共同作用对肌动球蛋白解离影响的Western blotting转印图(A)和定量分析图(B)
Fig.6 Western blotting (A) and quantitative analysis (B) on the effects of Ca
2+alone or in combination with ATP, ADP or AMP on actomyosin dissociation
由图6A可知,Ca 2+和ATP共同作用于肌动球蛋白,能观察到大量肌动蛋白的生成。Ca 2+和AMP共同作用于肌动球蛋白时,亦可观察到肌动球蛋白发生解离。由图6B可知,Ca 2+和ATP共同处理组的肌动蛋白条带灰度较其他处理组和对照组显著增加(P<0.05);Ca 2+和AMP共同处理组的肌动蛋白条带灰度虽然有增加,但与Ca 2+和ADP共同处理组和对照组相比则无显著差异(P>0.05)。
动物宰杀后,机体会在一段时间后发生僵直,大量的肌动蛋白和肌球蛋白结合生成肌动球蛋白,并逐渐由较弱的结合状态变成较强的结合状态,使嫩度降低 [2]。Okitani等 [7]研究发现将鸡肉、猪肉、牛肉等在65 ℃加热条件下,均能观察到明显的肌动球蛋白解离现象以及肌动蛋白含量的增多,并且有较好的嫩度。Li Shengjie等 [15]研究认为,肌动蛋白和肌球蛋白之间相互结合力的减弱、肌节长度的增加,能够导致肌肉微观结构的瓦解。因此,探讨肌动球蛋白的解离因素、并使肌动球蛋白大量的解离以提高肉品嫩度的研究具有重要意义。
目前,普遍认为肌原纤维紧密结构弱化是肉品嫩度改善的主要原因 [16-18],而Ca 2+在弱化肌原纤维完整结构的过程中起到了重要的作用。目前,关于Ca 2+的作用机理主要有两个方面:一是认为Ca 2+可以激活钙蛋白酶,对肌原纤维蛋白进行降解 [19-21];二是钙理论 [23-24],Takahashi等 [24]研究发现将钙蛋白酶抑制后,单独的Ca 2+亦能作用于Z盘,使之崩溃、断裂,因此认为Ca 2+才是弱化肌原纤维骨架的主要原因,而非钙蛋白酶的作用。本实验中,在Ca 2+浓度为7.5~64 mmol/L条件下,发现对肌动球蛋白的解离无促进作用,反而使肌动蛋白含量显著降低,可能原因是在外源较高浓度Ca 2+的作用下,肉中的μ-钙蛋白酶发生自溶,使得酶活力下降,反而促进更多的肌动蛋白和肌球蛋白结合;当Ca 2+浓度升高到200 mmol/L时,观察到肌动蛋白含量显著性增加,即大量的肌动球蛋白发生解离,此现象与Takahashi等 [24]所认为的钙理论相符,即在高浓度的Ca 2+作用下肌动球蛋白发生降解。
Okitani等 [12]研究认为AMP对肌动球蛋白的解离有促进作用,本实验得到与此较为相似的结果,8 mmol/L AMP可使肌动球蛋白发生显著性的解离效果,随着AMP浓度的提高,肌动球蛋白的解离与8 mmol/L AMP处理组相比无显著性差异。此外,ADP对肌动球蛋白的解离也有显著效果。但ATP对肌动球蛋白的解离无促进作用,此结论与Benjakul等 [13]研究单独的ATP对肌动球蛋白的解离相似。
Benjakul等 [13]研究发现,用5 mmol/L焦磷酸盐与5 mmol/L或10 mmol/L MgCl 2共同作用于肌动球蛋白时,能显著地促进肌动球蛋白的解离;而当ATP与MgCl 2共同作用于肌动球蛋白时,未发现对肌动球蛋白的解离有促进作用。本实验用16 mmol/L Ca 2+与16 mmol/L ATP共同作用于肌动球蛋白时,可以观察到肌动蛋白含量显著增加,表明有大量的肌动球蛋白发生解离,而ADP和Ca 2+共同作用肌动球蛋白时,肌动蛋白含量未增加;AMP与Ca 2+共同作用于肌动球蛋白时,虽有观察到肌动蛋白含量增加,但较ATP和Ca 2+处理组相比其促进作用要弱很多。可能的原因是,较高浓度的外源Ca 2+先对肌原纤维骨架蛋白发生作用,弱化它们之间的作用力,然后在ATP大量供能条件下,肌动球蛋白发生解离。
细胞内发生着复杂的生化反应,影响肌动球蛋白解离的因素远非目前所了解的这些,可能有更多种类的内源小分子物质影响着肌动球蛋白的解离,而这些小分子物质及细胞环境的变化等因素是如何共同影响肌动蛋白和肌球蛋白的结合状态及结合程度,目前还无明确的解释,因此,还需要做更进一步的研究,以对嫩度的变化机理有更加深入的了解,进而改善肉的嫩度、提高肉的品质。
参考文献:
[1] SHACKELFORD S D, WHEELER T L, MEADE M K, et al. Consumer impressions of tender select beef[J]. Journal of Animal Science, 2001, 79(10): 2605-2614.
[2] 李胜杰, 徐幸莲, 周光宏. 宰后肌动球蛋白解离对肉品嫩度的影响研究进展[J]. 食品科学, 2010, 31(21): 442-445.
[3] LEE Y S, SAHA A, XIONG R, et al. Changes in broiler breast fillet tenderness, water-holding capacity, and color attributes during long-term frozen storage[J]. Journal of Food Science, 2008, 73(4): E162-E168.
[4] TOMBERG E. Effects of heat on meat proteins-implications on structure and quality of meat products[J]. Meat Science, 2005, 70(3): 493-508.[5] TAKAHASHI K, NAKAMURA F, INOUE A. Postmortem changes in the actin-myosin interaction of rabbit skeletal muscle[J]. Journal of Biochemistry, 1981, 89(1): 321-324.
[6] KOOHMARAIE M. Biochemical factors regulating the toughening and tenderization processes of meat[J]. Meat Science, 1996, 43(1): 193-201.[7] OKITANI A, ICHINOSE N, ITOH J, et al. Liberation of actin from actomyosin in meats heated to 65℃[J]. Meat Science, 2009, 81(3): 446-450.
[8] GEESINK G H, TAYLOR R G, BEKHIT A, et al. Evidence against the non-enzymatic calcium theory of tenderization[J]. Meat Science,2001, 59(4): 417-422.
[9] ILIAN M A, MORTON J D, KENT M P, et al. Intermuscular variation in tenderness: association with the ubiquitous and muscle-specific calpains[J]. Journal of Animal Science, 2001, 79(1): 122-132.
[10] 黄明, 赵莲, 徐幸莲. 钙离子和钙激活酶外源抑制剂对牛肉钙激活酶活性和超微结构的影响[J]. 南京农业大学学报, 2004, 27(4): 101-104.
[11] YANG R, OKITANI A, FUJIMAKI M. Studies on myofibrils from the stored muscle. Part I. Post-mortem changes in adenosine triphosphatase activity of myofibrils from rabbit muscle[J]. Agricultural Biological Chemistry, 1970, 34(12): 1765-1772.
[12] OKITANI A, ICHINOSE N, KOZA M, et al. AMP and IMP dissociate actomyosin into actin and myosin[J]. Bioscience, Biotechnology and Biochemistry, 2008, 72(8): 2005-2011.
[13] BENJAKUL S, VISESSANGUAN W, AEWSIRI T, et al. Dissociation of natural actomyosin from kuruma prawn muscle induced by pyrophosphate[J]. Food Chemistry, 2007, 102(1): 295-301.
[14] 董晗, 王道营, 张牧焓. 不同加热温度对鸭肉肌动球蛋白解离的影响[J]. 食品工业科技, 2012, 33(20): 120-124.
[15] LI Shengjie, XU Xinglian, ZHOU Guanghong. The roles of actinmyosin interaction and proteolysis in tenderization during the aging of chicken muscle[J]. Poultry Science, 2012, 91(1): 150-160.
[16] NISHIMURA T. The role of intramuscular connective tissue in meat texture[J]. Animal Science Journal, 2010, 81(1): 21-27.
[17] HUFF LONERGAN E, ZHANG W G, LONERGAN S M. Biochemistry of postmortem muscle-lessons on mechanisms of meat tenderization[J]. Meat Science, 2010, 86(1): 184-195.
[18] SIKES A, TOMBERG E, TUME R. A proposed mechanism of tenderising post-rigor beef using high pressure-heat treatment[J]. Meat Science, 2010, 84(3): 390-399.
[19] 黄明, 黄峰, 黄继超. 内源性蛋白酶对宰后肌肉嫩化机制研究进展[J].中国农业科学, 2011, 44(15): 3214-3222.
[20] KOOHMARAIE M, BABIKER A S, SCHROEDER A L, et al. Acceleration of postmortem tenderization in ovine carcasses through activation of Ca 2+-dependent proteases[J]. Journal of Food Science,1988, 53(6): 1638-1641.
[21] KOOHMARAIE M, GEESINK G H. Contribution of postmortem muscle biochemistry to the delivery of consistent meat quality with particular focus on the calpain system[J]. Meat Science, 2006, 74(1): 34-43.
[22] HATTORI A, TAKAHASHI K. Calcium-induced weakening of skeletal muscle Z-disks[J]. Journal of Biochemistry, 1982, 92(2): 381-390.
[23] TAKAHASHI K, NAKAMURA F, OKAMOTO M. A myofibrillar component that modifies the actin-myosin interaction in postrigor skeletal muscle[J]. Journal of Biochemistry, 1982, 92(3): 809-815.
[24] TAKAHASHI K, KIM O H, KOJI Y. Calcium-induced weakening of Z-disks in postmortem skeletal muscle[J]. Journal of Biochemistry,1987, 101(3): 767-773.
Actomyosin Dissociation as Influenced by Ca 2+, ATP, ADP and AMP
DENG Shaoying
1,2, WANG Daoying
1,*, ZHANG Muhan
1, BIAN Huan
1, WU Haihong
1, ZHU Yongzhi
1, GENG Zhiming
1, LIU Fang
1, XU Weimin
1
(1. Institute of Agricultural Products Processing, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China; 2. Key Laboratory of Meat Processing and Quality Control, Ministry of Education, Nanjing Agricultural University, Nanjing 210095, China)
Abstract:To understand the potential factors that promote the dissociation of actomyosin, actomyosin was extracted from duck breast muscle and the effect of Ca 2+, ATP, and its degradation products on actomyosin dissociation was investigated. The dissociation of actomyosin was evaluated by measuring changes in actin content by Western blotting analysis. Results showed that there was no significant change in actomyosin dissociation when it was treated with 7.5-64 mmol/L Ca 2+(P >0.05), whereas actomyosin dissociation was enhanced significantly by 200 mmol/L Ca 2+treatment (P < 0.05). ATP treatment alone did not result in a significant change in actomyosin dissociation (P > 0.05), but combinatorial treatment with Ca 2+and ATP significantly increased actomyosin dissociation (P < 0.05). Both ADP or AMP remarkably promoted actomyosin dissociation (P < 0.05), but there was no significant difference among different concentration groups (P > 0.05). These results suggest that actomyosin dissociation is significantly improved when the actomyosin is treated with ADP, AMP, or combination of Ca 2+and ATP.
Key words:duck; actomyosin; Western blotting; dissociation; actin
中图分类号:TS251.68
文献标志码:A
文章编号:1002-6630(2015)03-0018-05
doi:10.7506/spkx1002-6630-201503004
收稿日期:2014-03-25
基金项目:国家自然科学基金青年科学基金项目(31101312)
作者简介:邓少颖(1992—),女,硕士研究生,研究方向为肉品质量与安全。E-mail:shaoyinglucky@163.com
*通信作者:王道营(1979—),男,副研究员,博士,研究方向为肉品加工与质量控制。E-mail:wdy0373@aliyun.com