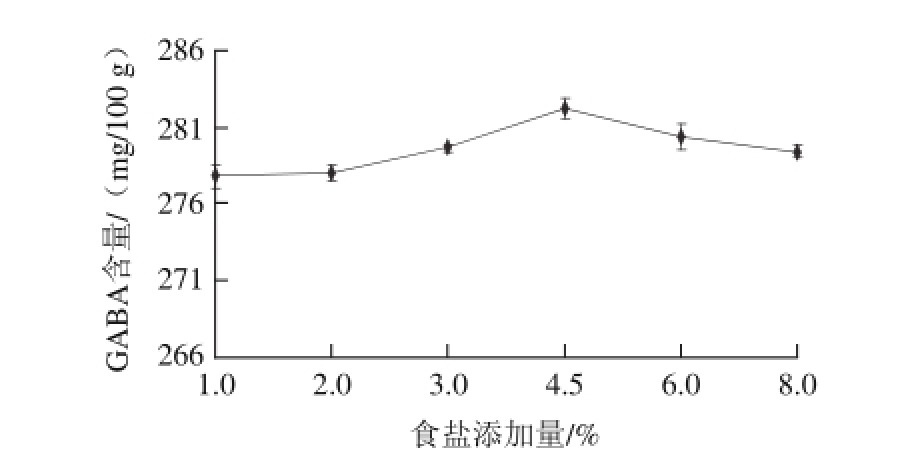
图1 不同食盐添加量处理对发酵酸肉GABA含量的影响
Fig.1 Impact of salt concentration on GABA amount in fermented pork
李成龙,袁 军,刘淑贞,周才琼*
(西南大学食品科学学院,重庆市特色食品工程技术研究中心,重庆 40071 5)
摘 要:以新鲜猪瘦肉为原料,研究米粉及食盐添加量对发酵60 d成品酸肉蛋白质降解及γ-氨基丁酸(γ-amino butyric acid,GABA)含量的影响,并利用响应面法对酸肉发酵工艺条件进行优化以获取高GABA含量的酸肉。结果表明:米粉和食盐添加量可影响蛋白质降解及GABA的形成,制备高GABA含量酸肉的最佳发酵工艺条件为米粉添加量12.73%、食盐添加量4.74%、发酵温度16.50 ℃。此时发酵60 d的成品酸肉中GABA含量最高为282.36 mg/100 g。
关键词:发酵酸肉;发酵条件;蛋白质降解;响应面法;γ-氨基丁酸
酸肉主要流行于贵州、四川、重庆、湖南等少数民族地区和汉族与少数民族杂居地区,是一种自然乳酸发酵的特色猪肉制品,至今已有两千多年的历史。相关研究表明 [1-7],经腌制发酵后的酸肉由于微生物生长繁殖,其蛋白质、总糖、脂肪、多肽氮及有机酸等含量变化,氨基酸组成改变,除了形成酸肉特有的清香、酯香和酱香风味外,其营养价值也发生了深刻变化。有报道从乳酸发酵产品中分离出可产生γ-氨基丁酸(γ-amino butyric acid,GABA)的乳酸菌 [8-10],作为现代营养学研究中重要的功能成分之一,GABA具有多种生物学功能,如抗高血压活性、免疫调节效应和抗氧化活性等 [11]。考虑酸肉厌氧发酵特点及发酵过程中乳酸菌大量生长繁殖,以及GABA富集的机理,本实验拟研究米粉添加量和食盐添加量对发酵60 d酸肉成品蛋白质降解及GABA形成的影响,为传统酸肉的功能化开发提供基础研究资料。
1.1 材料
新鲜猪瘦肉、大米 市售;大米粉,粳米淘洗后炒至微黄后磨成粉,过20 目筛。
1.2 试剂
NaCl、无水乙醇、次氯酸钠均为分析纯 成都市科龙化工试剂厂;K 2SO 4、CuSO 4、苯酚、硼酸等均为分析纯 重庆川东化工有限公司;GABA标准品 美国Sigma公司。
1.3 仪器与设备
722型可见光分光光度计 上海精密科学仪器有限公司;RE-52旋转蒸发器 上海亚荣生化仪器厂;Avant J-3DI型离心机 美国Beckman Coulter公司;101-1A型恒温鼓风干燥箱 上海东星建材实验设备有限公司;KjelFlex-360全自动凯氏定氮仪 北京德泉兴业商贸有限公司沈阳分公司。
1.4 方法
1.4.1 酸肉腌制工艺流程
原料肉→清洗→切成厚度约4 mm片状→加调味料(盐、米粉等)→揉制装坛→一定温度下密封发酵60 d→成品
1.4.2 不同食盐添加量对发酵60 d成品酸肉的影响
以米粉添加量12%、食盐添加量分别为1%、2%、3%、4.5%、6%和8%的条件下进行揉制装坛,置于15 ℃条件下密封发酵60 d。
1.4.3 不同米粉添加量对发酵60 d成品酸肉的影响
以食盐添加量4.5%,米粉添加量分别为2%、4%、8%、12%、16%和20%的条件下进行揉制装坛,置于15 ℃条件下密封发酵60 d。
1.4.4 响应面优化设计
根据1.4.2、1.4.3节试验结果选出食盐添加量和米粉添加量,并参考文献[12]选择发酵温度作为主要影响因素,进行响应面优化试验设计,进行Box-Behnken试验 [13]。
1.4.5 分析方法
1.4.5.1 测试样品前处理
取发酵60 d的酸肉,刷掉表面黏附的米粉、剔除可见的脂肪和筋膜。
1.4.5.2 粗蛋白和游离氨基酸含量测定
粗蛋白含量:按照GB/T 5009.5—2010《食品安全国家标准 食品中蛋白质的测定》;游离氨基酸含量:采用GB/T 8314—2002《茶 游离氨基酸总量测定》中的茚三酮比色法测定。
1.4.5.3 肌浆蛋白、肌原纤维蛋白、非蛋白氮和多肽氮
采用张亚军等 [14]的方法:微量凯氏定氮法。
1.4.5.4 GABA含量的测定 [15]
精确称取5.000 0 g切碎样品,磨细匀浆,置于150 mL圆底烧瓶中,加入体积分数60%乙醇溶液20 mL,然后置于88 ℃水浴回流萃取2 h后过滤,滤渣用10 mL 60%乙醇溶液分2 次洗涤,然后将3 次滤液合并于离心管中,3 000 r/min离心20 min,将离心管中上清液转入小烧杯内,置沸水浴蒸干,用8 mL蒸馏水充分溶解蒸干后的残渣并转入离心管,3 000 r/min离心20 min,取上清液备用。吸取处理后的样液0.5 mL于试管中,加入浓度为0.2 mol/L的硼酸缓冲液1.0 mL和质量分数为6%的苯酚1.0 mL,混匀后加入0.6 mL次氯酸钠溶液(有效氯≥5.1%),振荡。于沸水浴中加热10 min置于冰浴中20 min,待溶液出现蓝色后,加入2.0 mL 60%的乙醇,于吸收波长645 nm处测定吸光度,通过GABA质量浓度标准曲线,得到GABA的回归方程Y=1.999 6X+0.063 7(R 2=0.994 4),从而计算得到GABA含量。
1.5 数据分析
采用SPSS 19.0软件进行数据分析,采用Design Expert 7.0软件对响应面试验数据进行分析,运用Excel软件作图。
2.1 不同食盐添加量和米粉添加量对发酵60 d酸肉GABA含量的影响
2.1.1 不同食盐添加量对发酵60 d酸肉GABA含量的影响
图1 不同食盐添加量处理对发酵酸肉GABA含量的影响
Fig.1 Impact of salt concentration on GABA amount in fermented pork
猪瘦肉按照1.4.2节的方法分别添加12%的米粉和不同质量分数的食盐后在15 ℃条件下密封发酵,并测定发酵60 d成品酸肉GABA含量,结果如图1。GABA含量随食盐添加量的增加呈先增后降趋势,在277.84~282.30 mg/100 g之间,食盐添加量为4.5%时GABA含量最高,显著高于其他各处理组(P<0.05)。这可能与微生物的生长繁殖和蛋白质的降解有关,GABA是由谷氨酸在谷氨酸脱羧酶的作用下产生的,在食盐添加量低时,食盐添加量的增加,破坏原料肉蛋白质的结构,使蛋白酶更易作用于蛋白,加快蛋白质的降解,游离氨基酸增加,但食盐添加量过高则抑制了蛋白分解酶和微生物的活性,影响乳酸菌谷氨酸脱羧酶的产生。
2.1.2 不同米粉添加量对发酵60 d酸肉GABA含量的影响
猪瘦肉按照1.4.3节的方法分别添加4.5%的食盐和不同质量分数的米粉后在15 ℃条件下密封发酵,并测定发酵60 d成品酸肉中GABA含量,结果如图2。GABA含量随米粉添加量的增加呈先增后降趋势,在277.50~282.30 mg/100 g之间,米粉添加量为12%时酸肉GABA含量最高,显著高于米粉添加量分别为2%、4%、8%的处理(P<0.05),与米粉添加量为16%时酸肉中GABA含量之间无显著性差异(P>0.05)。可能微生物对于作为碳源的米粉存在饱和现象,外源米粉的添加量超过一定比例时,发酵酸肉原料中蛋白质含量下降较多,导致产品中GABA含量开始下降。
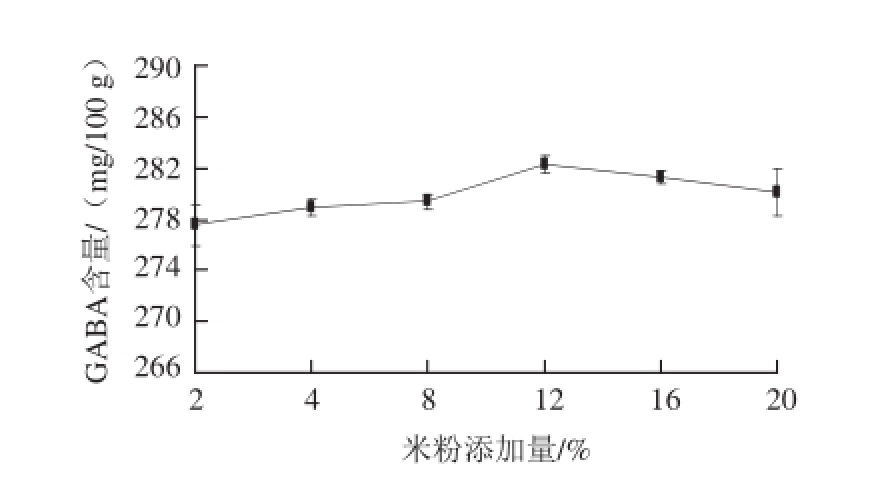
图2 不同米粉添加量处理对发酵酸肉GABA含量的影响
Fig.2 Impact of rice flour amount on GABA amount in fermented pork
2.2 不同食盐添加量和米粉添加量对发酵60 d酸肉蛋白质降解的影响
2.2.1 不同食盐添加量对发酵60 d酸肉蛋白质及降解的影响
猪瘦肉按照1.4.2节的方法分别添加12%的米粉和不同质量分数的食盐后在15 ℃条件下密封发酵,并测定发酵60 d成品酸肉蛋白质及降解产物含量变化,结果见表1,酸肉各蛋白含量均有所下降,而蛋白降解产物则有升高趋势,在食盐添加量4.5%时蛋白质含量降到最低。各蛋白和蛋白降解产物变化与GABA含量变化显著相关(P<0.05),其中肌浆蛋白和肌原纤维蛋白含量最高分别下降了2.07 g/100 g和2.28 g/100 g,非蛋白氮、多肽氮和游离氨基酸最高分别增加了0.61、0.41 g/100 g和0.41 g/100 g。考虑发酵中酸肉含水量的变化,以100 g干基计,则肌浆蛋白和肌原纤维蛋白含量最高分别下降了5.28 g和5.54 g,非蛋白氮、多肽氮和游离氨基酸最高分别增加了0.94、0.79 g和0.90 g。
表1 不同食盐添加量对发酵酸肉蛋白质降解的影响
Table1 Impact of salt concentration on protein degradation in fermented pork g/100 g
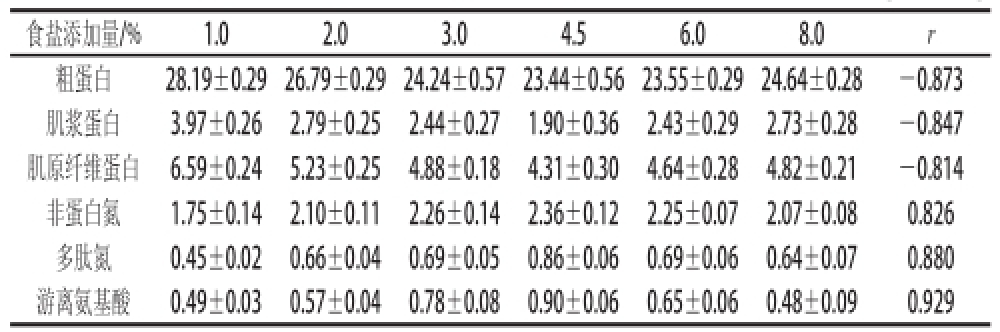
食盐添加量/%1.02.03.04.56.08.0r粗蛋白28.19±0.29 26.79±0.29 24.24±0.57 23.44±0.56 23.55±0.29 24.64±0.28-0.873肌浆蛋白3.97±0.262.79±0.252.44±0.271.90±0.362.43±0.292.73±0.28-0.847肌原纤维蛋白6.59±0.245.23±0.254.88±0.184.31±0.304.64±0.284.82±0.21-0.814非蛋白氮1.75±0.142.10±0.112.26±0.142.36±0.122.25±0.072.07±0.080.826多肽氮0.45±0.020.66±0.040.69±0.050.86±0.060.69±0.060.64±0.070.880游离氨基酸0.49±0.030.57±0.040.78±0.080.90±0.060.65±0.060.48±0.090.929
2.2.2 不同米粉添加量对发酵60 d酸肉蛋白质降解的影响
猪瘦肉按1.4.3节方法分别添加4.5%的食盐和不同质量分数的米粉后在15 ℃条件下密封发酵,并测定发酵60 d成品酸肉中蛋白质及降解产物。结果见表2,酸肉各蛋白质含量均有所下降,而蛋白降解产物则有升高趋势,在米粉添加量12%时蛋白质降解达到最大。各蛋白和蛋白降解产物变化与GABA含量变化显著相关(P<0.05),其中肌浆蛋白、肌原纤维蛋白含量最高分别下降了3.06 g和3.08 g,非蛋白氮、多肽氮和游离氨基酸最高分别增加了0.97、0.40 g和0.51 g。考虑到发酵中酸肉含水量的差异,以100 g干基计,则肌浆蛋白和肌原纤维蛋白含量最高分别下降了9.10 g和10.39 g,非蛋白氮、多肽氮和游离氨基酸最高分别增加了1.38、0.63 g和0.89 g。酸肉发酵中蛋白质的降低主要来自肌浆蛋白和肌原纤维蛋白的降解,多肽氮、非蛋白氮和游离氨基酸的增加对发酵酸肉形成特有的风味或特定功能成分如GABA有利。
表2 不同米粉添加量处理对发酵酸肉蛋白质降解的影响
Table2 Impact of rice flour content on protein degradation in fermented pork g/100 g
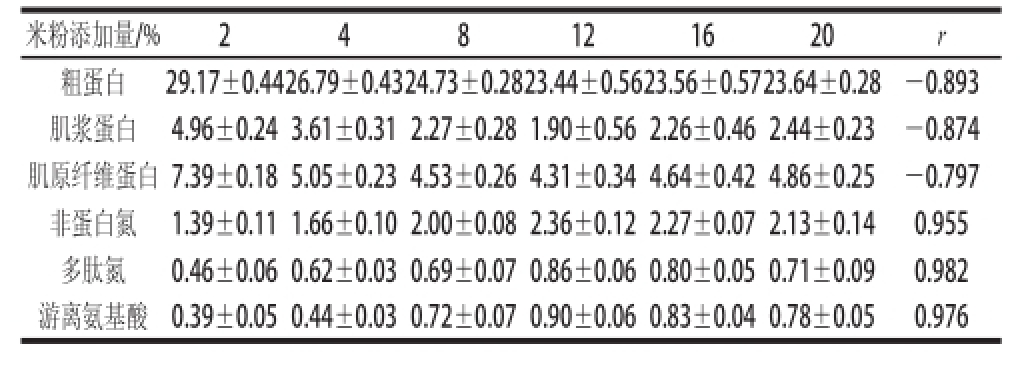
米粉添加量/%248121620r粗蛋白29.17±0.4426.79±0.4324.73±0.2823.44±0.5623.56±0.5723.64±0.28-0.893肌浆蛋白4.96±0.24 3.61±0.31 2.27±0.28 1.90±0.56 2.26±0.46 2.44±0.23-0.874肌原纤维蛋白7.39±0.18 5.05±0.23 4.53±0.26 4.31±0.34 4.64±0.42 4.86±0.25-0.797非蛋白氮1.39±0.11 1.66±0.10 2.00±0.08 2.36±0.12 2.27±0.07 2.13±0.140.955多肽氮0.46±0.06 0.62±0.03 0.69±0.07 0.86±0.06 0.80±0.05 0.71±0.090.982游离氨基酸0.39±0.05 0.44±0.03 0.72±0.07 0.90±0.06 0.83±0.04 0.78±0.050.976
2.3 响应面法优化对发酵酸肉GABA富集的影响
以上述单因素试验为基础,选择米粉添加量、食盐添加量、发酵温度进行三因素三水平Box-Behnken响应面试验设计,以发酵60 d成品酸肉中GABA含量为指标,通过Design Expert7.0软件对试验数据进行回归分析,预测发酵条件对GABA含量的最佳工艺参数。试验结果见表3,回归分析见表4。
表3 响应面试验设计方案及结果
Table3 Response surface experimental design and results

试验号A米粉添加量/% Y GABA含量/(mg/100 g)11(16)0(4.5)1(25)281.26±0.62 20(12)00(15)282.63±0.13 3 11(6.0)0280.39±0.46 4 0 1-1(5)280.01±0.38 5 0 0 0282.43±0.15 6-1(8)01279.84±0.34 7 1 0 -1280.01±0.45 B食盐添加量/% C发酵温度/℃0282.23±0.23 9 0-1(3.0)1279.79±0.18 101-10280.21±0.56 11-1-10279.86±0.46 12000282.35±0.15 130-1-1279.59±0.08 14-110280.39±0.14 15000282.23±0.28 16011280.76±0.62 17-10-1279.42±0.13 8 0 0
经分析得到发酵条件与GABA含量之间的二次多项式模型为:Y=282.37+0.30A+0.26B+0.33C-0.088AB+ 0.21AC+0.14BC-1.03A 2-1.13B 2-1.21C 2。
对上述回归模型进行方差分析,结果如表4。该模型P=0.000 1<0.01,说明方差模型差异显著;模型失拟项P=0.064 3>0.05,即模型失拟项差异不显著,模型选择合适 [16-17]。模型决定系数R 2=0.972 3,即由3 个影响因素引起GABA含量的变异为97.23%,说明此模型相关性良好,可用此方程对不同发酵条件下酸肉中GABA含量进行预测。由表4可知,因素A、B、C对酸肉中GABA含量影响显著(P<0.05),AB、AC、BC对酸肉中GABA含量影响不显著(P>0.05),A 2、B 2、C 2对酸肉中GABA含量的影响极显著(P<0.01)。
表4 以GABA含量为响应值的回归模型方差分析
Table4 Analysis of variance of regression model with GABA amount as response value

注:R 2 Adj=0.936 6;**. P<0.01,表示差异极显著;*. P<0.05,表示差异显著。
方差来源均方和自由度均方F值P值显著性模型20.2692.2527.270.000 1** A0.7010.708.430.022 8* B0.5510.556.680.036 2* C0.8610.8610.400.014 6* AB0.03110.0310.370.561 7不显著AC0.1710.172.090.191 8不显著BC0.07610.0760.920.370 3不显著A 24.5014.5054.460.000 2** B 25.3615.3664.94<0.000 1** C 26.1516.1574.47<0.000 1**残差0.5870.083失拟项0.4730.165.620.064 3不显著纯误差0.1140.028总和20.8416
当发酵温度为15 ℃时,米粉添加量和食盐添加量交互作用对GABA含量的影响如图3。从响应面曲线可知,米粉添加量和食盐添加量对酸肉GABA含量影响的交互作用不显著(P>0.05)。随米粉添加量和食盐添加量增加,GABA含量呈先增后减趋势,在米粉添加量12%左右,食盐添加量4.5%左右时,GABA含量达最大值。


图3 米粉添加量和食盐添加量交互作用对GABA含量影响的响应面和等高线图
Fig.3 Response surface and contour plots for the effect of rice flour and salt content on GABA amount
当食盐添加量为4.5%时,米粉添加量和发酵温度交互作用对GABA含量的影响如图4。米粉添加量和发酵温度对酸肉中GABA含量影响的交互作用不显著(P>0.05)。随米粉添加量的增加和发酵温度升高,GABA含量呈先增后减的趋势,米粉添加量在12%左右,发酵温度15 ℃左右时,GABA含量达最大值。

图4 米粉添加量和发酵温度交互作用对GABA含量影响的响应面和等高线图
Fig.4 Response surface and contour plots for the effect of rice flour amount and fermentation temperature on GABA amount
当米粉添加量为12%时,食盐添加量和发酵温度交互作用对GABA含量的影响如图5。食盐添加量和发酵温度对酸肉中GABA含量影响的交互作用不显著(P>0.05)。随食盐添加量和发酵温度升高,GABA含量呈先增后减趋势,食盐添加量在4.5%左右,发酵温度15 ℃左右时,GABA含量达最大值。
根据Box-Behnken试验所得结果和二次多项回归方程,利用Design Expert软件获得了发酵60 d成品酸肉GABA含量最高时各因素最佳条件为:米粉添加量12.73%、食盐添加量4.74%、发酵温度16.50 ℃。在此条件下,GABA含量预测值282.44 mg/100 g。为检验模型预测准确性,在此工艺条件下测定发酵60 d酸肉GABA含量为282.36 mg/100 g,与预测值接近。表明优化得到的富集工艺参数准确可靠,模型合适有效,有实践参考价值。
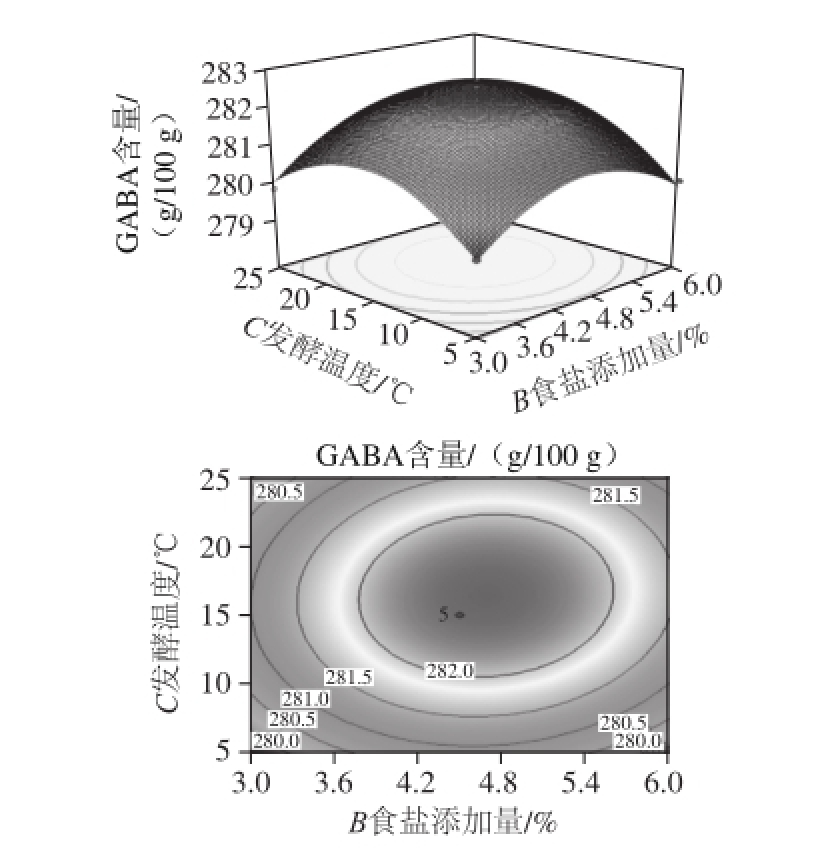
图5 食盐添加量和发酵温度交互作用对GABA含量影响的响应面和等高线图
Fig.5 Response surface and contour plots for the effect of salt amount and fermentation temperature on GABA amount
渝黔地区传统发酵酸肉为自然乳酸发酵食品,有大量微生物特别是乳酸菌的生长,乳酸菌优势菌群主要是片球菌属(Pediococcus)和乳杆菌属(Lactobacillus) [5]。相关研究显示乳酸菌、酵母菌和微球菌等可分泌胞外酶 [18-19],微生物胞外蛋白酶可降解蛋白质产生活性肽类及游离氨基酸 [20],在蛋白质降解中也可形成一定的挥发性盐基氮,已有报道显示发酵60 d的酸肉瘦肉的挥发性盐基氮为19.56 mg/100 g [6],符合国家标准中的二级鲜度15~25 mg/100 g。酸肉发酵中米粉添加量、食盐添加量及发酵温度都会影响微生物的生长繁殖,进而影响蛋白质的降解和GABA的形成。米粉为乳酸菌等微生物的生长及分解活动提供碳源,随碳源增加,微生物活动增加,微生物胞外蛋白酶导致蛋白质降解、非蛋白氮增加和GABA形成;当碳源饱和时其影响变小。食盐添加量变化会影响乳酸菌生长,从而影响谷氨酸转化成GABA。
GABA是由谷氨酸在谷氨酸脱羧酶作用下产生,谷氨酸由蛋白质和多肽降解而来,酸肉中乳酸菌生长产生胞外酶如谷氨酸脱羧酶可作用于谷氨酸产生GABA,推测低质量分数食盐可破坏原料肉蛋白质结构,使蛋白酶更易作用于蛋白,利于谷氨酸产生,高质量分数食盐对乳酸菌生长有一定抑制,影响乳酸菌胞外酶的生产。食盐不仅对谷氨酸脱羧酶有弱抑制作用,还能增加谷氨酸脱羧酶的胶体体系稳定性,二者共同作用降低谷氨酸脱羧酶活性,不利于GABA积累 [21]。
本实验结果表明米粉添加量、食盐添加量和发酵温度均对发酵60 d酸肉中GABA含量有显著影响(P<0.05),酸肉发酵工艺优化条件为米粉添加量12.73%、食盐添加量4.74%、发酵温度16.50 ℃时,发酵60 d的成品酸肉中GABA含量最高。
参考文献:
[1] 李宗军, 江汉湖. 中国传统酸肉发酵过程中微生物的消长变化[J].微生物学通报, 2004, 31(4): 9-13.
[2] 李华丽, 何煜波. 酸肉生产主发酵期发酵条件的确定[J]. 中国食物与营养, 2005(4): 40-43.
[3] 章德法, 徐为民, 徐幸莲. 侗族发酵酸肉中乳酸菌的筛选及鉴定[J].江苏农业学报, 2009, 24(6): 986-988.
[4] 娄爱华, 邹畅, 李华丽. 酸肉生产主发酵期发酵条件的确定[J]. 肉品卫生, 2005(6): 20-25.
[5] 李艳芳. 渝黔地区传统发酵酸肉的理化特性及优势微生物区系研究[D]. 重庆: 西南大学, 2007.
[6] 陈东华. 酸腌肉的营养及安全性研究[D]. 重庆: 西南大学, 2008.
[7] 代小容. 酸肉腌制过程中风味形成研究[D]. 重庆: 西南大学, 2009.
[8] KOMATSUZAKI N, SHIMA J, KAWAMOTO S, et al. Production of γ-aminobutyric acid (GABA) by Lactobacillus paracasei isolated from traditional fermented foods[J]. Food Microbiology, 2005, 22(6): 497-504.
[9] CHOI J, SABIKHI L, HASSAN A, et al. Bioactive peptides in dairy products[J]. International Journal of Dairy Technology, 2012, 65(1):1-12.
[10] 穆琳, 阮晖, 唐彦捷, 等. 产γ-氨基丁酸乳酸菌的分离筛选及其主要性能[J]. 中国食品学报, 2009, 9(3): 20-25.
[11] 朱晓立, 裘晖. γ-氨基丁酸的生物学功能及其在食品上的应用[J]. 食品工程, 2008(2): 34-37.
[12] 李华丽, 何煜波. 酸肉生产前期发酵条件的确定[J]. 肉类工业,2005(2): 24-30.
[13] O'BRIEN S, WANG Y J. Susceptibility of annealed starches to hydrolysis by α-amylase and glucoamylase[J]. Carbohydrate Polymers,2008, 72(4): 597-607.
[14] 张亚军, 陈有亮. 金华火腿蛋白降解及其影响因素的研究[J]. 中国食品学报, 2006, 6(3): 16-24.
[15] 何秋云. 马铃薯γ-氨基丁酸(GABA)富集技术及其加工制品的工艺研究[D]. 南宁: 广西大学, 2011.
[16] DWEVEDI A, KAYASTHA A M. Optimal immobilization of β-galactosidase from Pea (PsBGAL) onto Sepha dex and chitosan beads using response surface methodology and its applications[J]. Bioresource Technology, 2009, 100(10): 2667-2675.
[17] RANGONESE R, MACKA M, HUGHES J, et al. The use of Box-Behnken expermental design in the optimisation and robustness testing of a capillary electrophoresis method for the analysis of ethambutol hydrochloride in a pharmaceutical formulation[J]. Journal of Pharmaceutical and Biomedical Analysis, 2006, 27(6): 995-1007.
[18] 陈超, 高学军, 刘营, 等. 产蛋白酶乳酸茵的筛选及蛋白酶的纯化[J].乳业科学与技术, 2008(4): 160-162.
[19] 江洁, 吴耘红, 蒋继丰, 等. 豆豉中微球菌的分离及其产蛋白酶特性研究[J]. 中国调味品, 2004, 29(5): 22-26.
[20] NIELSEN M S, MARTINUSSEN T, FLAMBARD B, et al. Peptide profiles and angiotensin-I-converting enzyme inhibitory activity of fermented milk products: effects of bacterial strain, fermentation pH,and storage time[J]. International Dairy Journal, 2009, 19: 155-165.
[21] 杨胜远, 陆兆新. 中性盐对谷氨酸脱羧酶活性的影响机制[J]. 分子化学, 2007, 21(6): 556-560.
Impact of Amounts of Rice Flour and Salt Added to Fermented Pork on Protein Degradation and γ-Amino Butyric Acid (GABA) Formation
LI Chenglong, YUAN Jun, LIU Shuzhen, ZHOU Caiqiong*
(Chongqing Special Food Engineering and Technology Research Center, College of Food Science, Southwest University,Chongqing 400715, China)
Abstract:The fermentation process of fresh lean pork with added rice fl our and salt to produce sour pork was optimized for higher content of γ-amino butyric acid (GABA) by response surface methodology. The effect of different amounts of rice fl our and salt on protein degradation and GABA content after 60 days of fermentation was assessed. Results indicated that the amounts of rice fl our amount and salt could affect the formation of GABA and protein degradation. The optimum fermentation conditions for sour pork were addition of 12.73% of rice fl our and 4.74% of salt and fermentation at 16.50 ℃. Under these optimized conditions, the maximum content of GABA was 282.36 mg/100 g.
Key words:fermented pork; fermentation conditions; protein degradation; response surface methodology; γ-amino butyric acid (GABA)
中图分类号:TS251.1
文献标志码:A
文章编号:1002-6630(2015)13-0202-05
doi:10.7506/spkx1002-6630-201513037
收稿日期:2014-09-01
作者简介:李成龙(1990—),男,硕士研究生,研究方向为食品化学与营养学。E-mail:654196032@qq.com
*通信作者:周才琼(1964—),女,教授,博士,研究方向为食品营养化学。E-mail:zhoucaiqiong@swu.edu.cn