表1 qRT-PCR引物序列
Table1 Primer sequences for qRT-PCR

?
赵 丹 1,苏 宁 2,杨 丽 2,郑洪艳 2,霍 彤 1,王昌涛 1,*
(1.北京工商大学理学院,北京 100048;2.中国检验检疫科学研究院,北京 100176)
摘 要:目的:通过体外实验方法从分子水平对蓝莓叶总黄酮的抗炎功效进行评价。方法:建立人永生化表皮细胞 HaCaT与蓝莓叶总黄酮的共培养体系,利用反转录实时荧光定量聚合酶链式反应检测在不同培养时间点( 0、 15、 30、 60、 120 min)白细胞介素 -6( interleukin-6, IL-6)、 IL-8、干扰素诱导蛋白 -10( interferon-inducible prot ein-10, IP-10)等炎症细胞因子相关基因表达情况,观察蓝莓叶总黄酮的添加对这些基因表达的影响,并探讨蓝莓叶总黄酮在分子水平上的抗炎机理。结果:蓝莓叶总黄酮的添加能够提高抗炎因子 IL-6、 CXCR-2 基因的表达量( P< 0.05),降低促炎因子 IL-8、 IP-10 基因的表达量( P< 0.05),对 CXCR-1 基因的表达呈复杂型调节。结论:蓝莓叶总黄酮通过对炎症相关因子表达量产生影响,从而减弱促炎症反应,增强抗炎症反应,最终发挥抗炎功效。
关键词:蓝莓叶总黄酮;炎症因子;反转录实时荧光定量聚合酶链式反应;抗炎
由于动物实验所存在的伦理问题以及国际上对动物福利的日益关注,发展新的体外实验方法用以减少、优化和替代( reduction, refinement, replacement, 3R理论)动物实验成为当前研究的热点。反转录实时荧光定量聚合酶链式反应( real-time reverse transcription quantitative polymerase chain reaction, RT-qPCR)技术因其具有特异性强、灵敏度高以及重复性好等优势,在动物替代实验领域发挥了日益重要的作用。此技术不仅避免了 PCR产物污染产生的假阳性结果和非特异性扩增问题,同时还能够对标本模板进行精确定量 [1-2]。
皮肤在受到外界药物刺激后会发生炎症反应。 在这个过程中,白细胞介素 -6( interleukin-6, IL-6)、 IL-8、干扰素诱导蛋白 -10( interferon-inducible protein-10, IP-10)等炎症因子和趋化因子的相关基因表达会受到影响。 IL-6能够刺激参与免疫反应的细胞增殖、分化并提高其功能活性; IL-8是巨噬细胞和上皮细胞等分泌的细胞因子,与其受体 CXCR-1和 CXCR-2结合而对中性粒细胞产生细胞趋化作用,从而实现对炎症反应的调节 [3-5]。
本实验利用反转录实时荧光定量 PCR技术,检测在人类永生化表皮细胞 HaCaT培养体系中蓝莓叶总黄酮的添加对这些细胞因子表达量的影响,从而进一步对这些细胞因子的表达与皮肤炎症反应的关系进行研究。
1.1材料与试剂
HaCaT细胞,购自上海斯信生物科技有限公司。
DMEM低糖培养基、二甲基亚砜 德国 Sigma公司;胎牛血清 美国 Gibco公司;青霉素 -链霉素双抗( 100×) 美国 Corning公司。
1.2仪器与设备
WJ-80A-Ⅱ CO 2培养箱 上海圣科仪器设备有限公司; ABI 7300型实时荧光定量 PCR仪、 GeneAmp PCR System 9700美国 ABI应用生物系统公司; Enduro水平电泳仪 美国 Labnet公司; 3-30K台式高速冷冻离心机、 1-14微型离心机 德国 Sigma公司; SW-CJ-1D超净工作台 上海启前电子科技有限公司;细胞培养瓶、细胞培养瓶板、细胞冻存管 美国 Corning/Costar公司。 1.3方法
1.3.1细胞培养及处理
从- 80℃冰箱中取出待用冻存的 HaCaT细胞,迅速置于 37℃水浴中解冻,在 1~ 2 min内轻微振动使其融化。 1 000 r/min离心 10 min,弃去上清液,在沉淀中加入 1 mL DMEM完全培养液,转移至 25 cm 2培养瓶中, 37℃、 5% CO 2培养。当细胞生长至 90%融合时,用 2 mL磷酸盐缓冲液( phosphate buffered saline, PBS)轻柔清洗两次,加入 200μL 0.25%胰酶后放入 CO 2培养箱中消化 5 min,放到显微镜下观察,当确认细胞从培养瓶壁上脱落后,加入 2 mL DMEM完全培养液终止消化, 800 r/min离心 10 min,弃去上清液,加入 4 mL培养液,进行细胞计数后,以 5× 10 5个 /cm 2的密度接种于 25 cm 2培养瓶中。
将消化后的 HaCaT细胞以 2× 10 6个 /孔的密度接种于 6孔板内,以 60μg /mL质量浓度蓝莓叶总黄酮处理细胞 0、 15、 30、 60、 120 min。
1.3.2细胞总 RNA提取与检测
待细胞密度长至 80%时,冰浴条件下,依次加入 1 mL Trizol、 0.2 mL氯仿,手摇振荡 2 min; 4℃、 8 000 r/min离心 15 min,取上层水相,加入等体积异丙醇(约 700μL),振荡,静置 15 min; 12 000 r/min离心 10 min,弃去上清液,向沉淀中加入 1 mL 70%乙醇洗涤, 8 000 r/min离心 10 min;重复进行 1次乙醇洗涤;室温条件下干燥 15 min,加入 40μL经焦碳酸二乙酯( diethyl pyrocarbonate, DEPC)处理的 ddH 2 O,溶解 10 min后于- 80℃保存 [6-7]。
琼脂糖凝胶电泳检测 RNA:将样品和 Marker加至凝胶上样孔, 180 V电压电泳 10 min。
1.3.3 cDNA第一链合成
使用 TINAGEN FastQuant RT Kit(含 gDNase) FastQuant cDNA第一链合成试剂盒(去基因组)进行 cDNA第一条链合成反应。
1.3.4反转录实时荧光定量 PCR分析
1.3.4.1引物设计
根据 NCBI中发布的IL-6、IL-8、IP-10等基因的序列,通过 Primer Express软件设计特异性引物,同时设计管家基因β-actin的特异性引物,各基因的引物序列见表 1。
表1 qRT-PCR引物序列
Table1 Primer sequences for qRT-PCR
?
1.3.4.2 反转录实时荧光定量 PCR反应
以 cDNA为模板,按照表 2配制 RT-qPCR反应体系(使用前,在 1 mL PrimeScript RT Enzyme MIX中加入 40μL的 ROX Reference Dye),用 ddH 2 O将反应体系补足到 20μL,分析IL-6、IL-8、IP-10等基因的表达量。
表2 反转录实时荧光定量PCR反应体系Table2 Real time fluorescence quantitative reverse transcription PCR
Table2 Real time f reaction system stem
试剂PrimeScript RTEnzyme MIX正向引物反向引物ddH 2OcDNA体积/μL10.40.50.57.61
2.1 HaCaT细胞培养
图 1为 HaCaT细胞培养贴壁前后形态比较,贴壁前 HaCaT细胞为规则的圆球体状,透明饱满,且可随培养液流动。当培养 2~ 4 h以后, HaCaT细胞开始呈现出片层状态贴壁生长,单个细胞形态不规则,其形态会因其所处培养空间大小的具体情况不同而变化;另一方面,随着培养液中营养物质的消耗和 HaCaT细胞自身生长过程中代谢物的产生,培养液的颜色(随 pH值变化)和细胞中内容物的颜色也与贴壁前的 HaCaT细胞有了显著的差别。这就决定了一定时间内需要根据培养瓶或者培养板中 HaCaT细胞生长的具体情况,而定期换新鲜培养液或者进行传代培养,才能保证 HaCaT细胞良好的生长状态。

图1 HaCaT细胞培养贴壁前(a)后(b)的形态(20×)
Fig.1 HaCaT cells before (a) and after (b) attached culture (20 ×)
HaCaT细胞对生长环境的要求相对其他动物细胞较为宽泛。 HaCaT细胞增殖速率快,培养 2~ 4 h即可贴壁, 36 h左右需要进行传代。细胞快速地增殖会引起培养液中营养物质在短时间内被大量消耗,限制了细胞进一步生长。因此,在考察蓝莓叶总黄酮对 HaCaT细胞的影响时,培养时间不宜过长。综合考虑,本实验中选取培养时间分别为 0、 15、 30、 60、 90、 120 min,在以上 6个时间点对添加蓝莓叶总黄酮的 HaCaT细胞生长状况进行考察。
2.2 HaCaT细胞总 RNA的提取
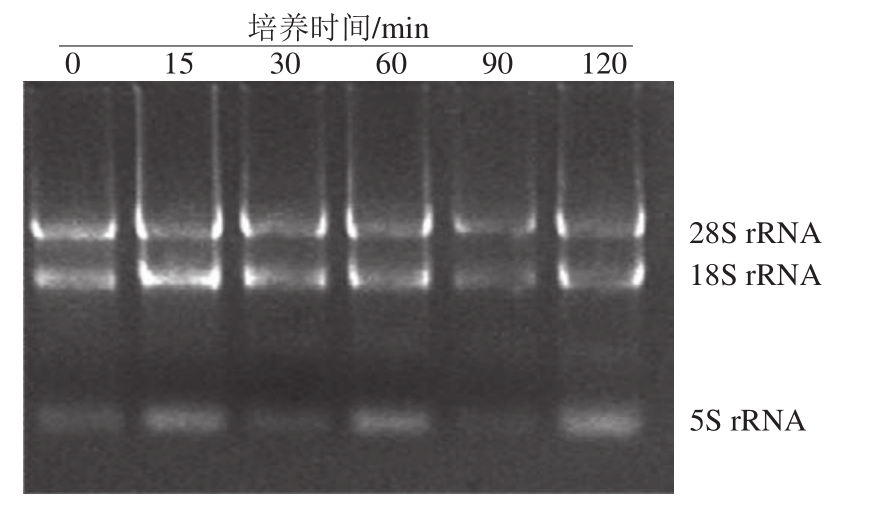
图2 HaCaT细胞总RNA琼脂糖凝胶电泳检测结果
Fig.2 Electrophoresis of total RNA from HaCaT cells
采用 1.0%琼脂糖凝胶电泳对提取的 HaCaT细胞 RNA进行完整性检测。如图 2所示, 28S rRNA与 18S rRNA带型清晰。另经紫外分光光度计测定,所提取 HaCaT细胞 RNA的A 260 nm /A 280 nm介于 1.8~ 2.0之间, RNA质量高,完整性好,适于反转录实时荧光定量 PCR研究 [8]。
2.3 RT-qPCR炎症因子特异性引物的检测
利用设计的β -actin、 IL-6和 IL-8的引物对目标产物进行检测,结果如图 3所示。以反转录得到的 cDNA为模板,通过设计的引物进行扩增,得到的产物利用 1%琼脂糖凝胶电泳进行检验,得到条带单一的、片段大小与预期目的条带大小一致的特异性引物,可进行下一步 RT-qPCR的实验。

图3 炎症因子IL-6、IL-8特异性引物检测结果
Fig.3 Electrophoresis of PCR amplified products with specific primers for β-actin, IL-6 and IL-8
2.4 RT-qPCR实验结果
以 HaCaT细胞为研究对象,考虑到其炎症因子相关基因表达量变化的时间大多在 2 h以内 [9],并且综合 HaCaT细胞生长情况,本实验选择 2 h以内作为蓝莓叶总黄酮处理 HaCaT细胞的考察时间。
2.4.1蓝莓叶总黄酮对 HaCaT细胞炎症因子类基因表达的影响
2.4.1.1蓝莓叶总黄酮对 HaCaT细胞IL-6表达的影响

图4 蓝莓叶总黄酮处理不同时间对HaCaT细胞IL--66表达量的影响
Fig.4 Influence of treatment time with blueberry leaf extracts on IL-6 expression
添加蓝莓叶总黄酮对 HaCaT细胞IL-6表达的影响如图 4所示,培养时间在 30 min内, HaCaT细胞的IL-6表达量相对于空白组略有下降;培养时间在 30~ 90 min内, HaCaT细胞的IL-6表达量明显升高;培养时间在 90~ 120 min内,IL-6表达量出现下降的趋势,回落到培养时间在 60 min时的表达水平,但远远高于空白组。整体来看,蓝莓叶总黄酮对 HaCaT细胞IL-6基因表达量的影响主要表现为在培养 30 min后的促进作用。对不同时间段IL-6表达量进行分析,发现与空白组相比,培养至 60、 90、 120 min时, HaCaT细胞IL-6表达量均显著或极显著升高(P< 0.05或P< 0.01)。 IL-6是一种在炎症反应和免疫反应中具有多种功能的细胞因子,其在炎症反应中可促进花生四烯酸的释放,刺激 T细胞活化和增殖,促进 B细胞中免疫球蛋白的生成,并诱导急性期反应蛋白的产生 [10]。蓝莓叶总黄酮对IL-6基因表达量的影响验证了蓝莓叶总黄酮通过影响花生四烯酸的代谢而起到抗炎作用。
2.4.1.2蓝莓叶总黄酮对 HaCaT细胞IL-8表达的影响
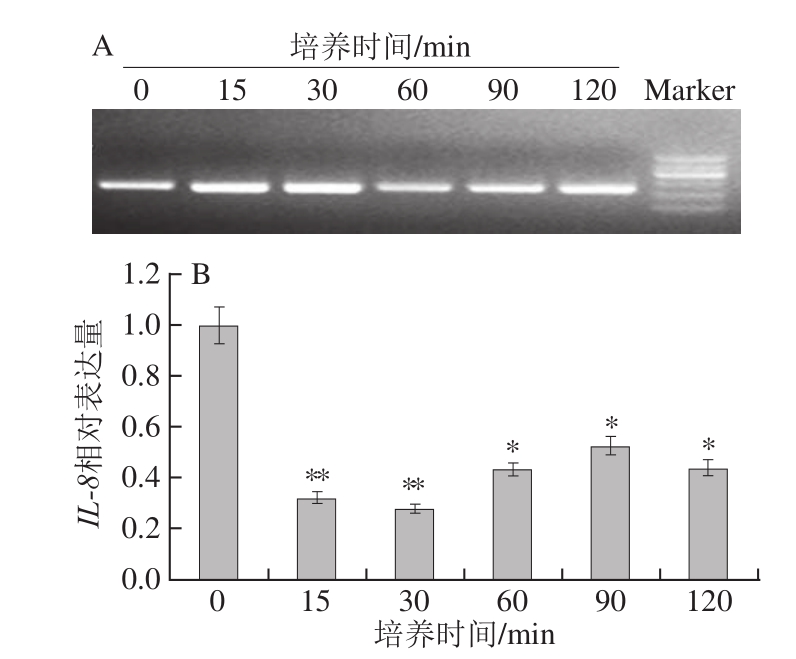
图5 蓝莓叶总黄酮处理不同时间对HaCaT细胞IL--88表达量的影响
Fig.5 Influence of treatment time with blueberry leaf extracts on IL-8 expression
蓝莓叶总黄酮对 HaCaT细胞IL-8表达量的影响如图 5所示,培养时间在 15 min内, HaCaT细胞IL-8表达量迅速下降,并在 30 min内保持缓慢下降的趋势,在 30~ 90 min内又缓慢上升,但远小于空白组,而在 90~ 120 min的培养时间内,IL-8表达量又有下降趋势。整体来看,蓝莓叶总黄酮对IL-8基因表达的影响主要体现在培养 0~ 15 min内迅速抑制其表达,并在 120 min的培养时间内总体保持抑制其表达的作用。对不同时间段IL-8表达量进行分析,发现与空白组相比,培养至 15、 30 min时, HaCaT细胞IL-8表达量极显著降低(P< 0.01),而培养至 60~ 120 min时,IL-8表达量稍有回升,但仍显著低于空白组(P< 0.05),这表明蓝莓叶总黄酮对 HaCaT细胞IL-8的表达具有抑制作用。IL-8在许多炎症中具有聚集和激活 T淋巴细胞、白细胞和血管内皮细胞的功能,局部产生的IL-8可以直接作用于表皮细胞,促进慢性炎症的发展 [11]。IL-8表达量的升高会使大量中性粒细胞向炎症区域聚集,释放炎症介质而加重炎症反应 [12-13]。蓝莓叶总黄酮抑制了IL-8的表达,从而间接地抑制了炎症的发生和发展。
2.4.2蓝莓叶总黄酮对 HaCaT细胞趋化因子相关基因表达的影响
2.4.2.1蓝莓叶总黄酮对 HaCaT细胞CXCR-1和CXCR-2表达的影响
CXCR-1和 CXCR-2是 CXCR家族中的两个重要成员,属 G蛋白偶联受体超家族成员。 CXCR-1和 CXCR-2会表达在正常的人内皮细胞表面,在急性炎症反应过程中,趋化因子能够促进内皮细胞释放大量促炎细胞因子 [14]。
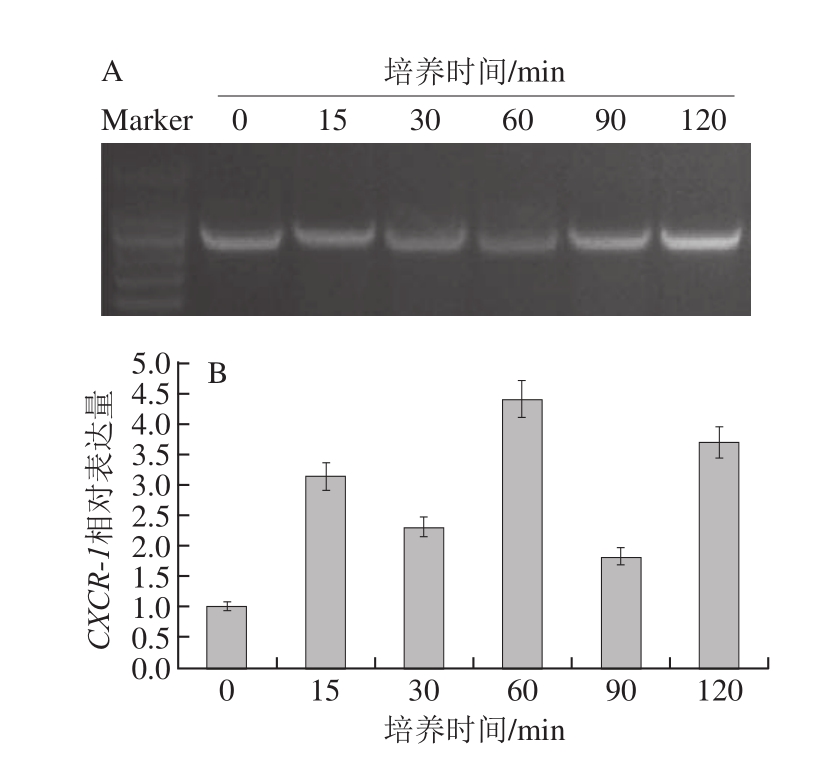
图6 蓝莓叶总黄酮处理不同时间对HaCaT细胞CXCR--11表达量的影响
Fig.6 Influence of treatment time with blueberry leaf extracts on CXCR-1 expression
如图 6所示,添加蓝莓叶总黄酮后, HaCaT细胞CXCR-1的表达量在培养 15 min内有明显升高的趋势,随后的培养过程中,CXCR-1的表达量被间隔性地促进和抑制,但总体水平都是高于空白组。对不同培养时间段 HaCaT细胞CXCR-1表达量进行统计学分析,发现与空白组相比,在培养的 120 min内, HaCaT细胞CXCR-1表达量的变化均不显著(P> 0.05)。以上结果表明,蓝莓叶总黄酮对 HaCaT细胞CXCR-1的表达有一定促进作用,但在促进和抑制间存在一定的动态平衡。
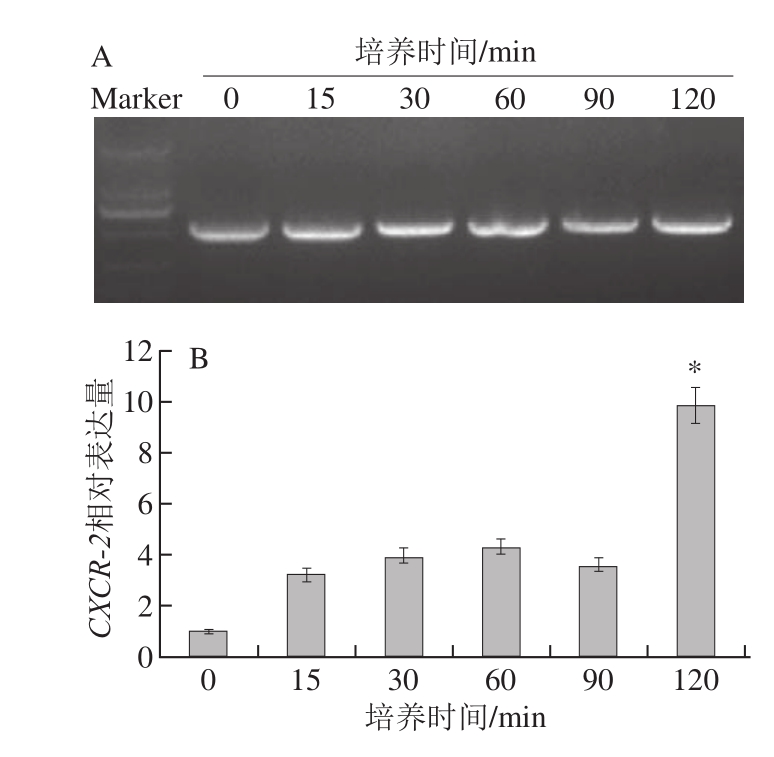
图7 蓝莓叶总黄酮处理不同时间对HaCaT细胞CXCR--22表达量的影响
Fig.7 Influence of treatment time with blueberry leaf extracts on CXCR-2 expression
如图 7所示,添加蓝莓叶总黄酮后, HaCaT细胞CXCR-2的表达量在培养 15 min内有平缓的升高,在随后培养的 15~ 90 min内,保持在较高的水平,而在培养的 90~ 120 min内,CXCR-2的表达量显著升高(P< 0.05)。以上结果表明,蓝莓叶总黄酮对 HaCaT细胞CXCR-2的表达是起促进作用的。
2.4.2.2蓝莓叶总黄酮对 HaCaT细胞IP-10表达的影响
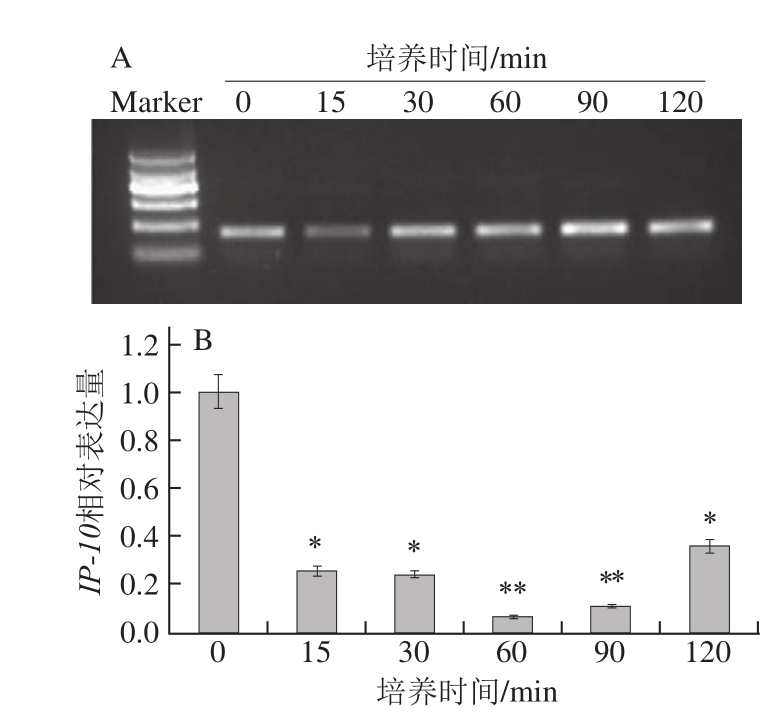
图8 蓝莓叶总黄酮处理不同时间对HaCaT细胞IP-10表达量的影响
Fig.8 Influence of treatment time by blueberry leaf extracts on IP-10 expression
如图 8所示,添加蓝莓叶总黄酮后, HaCaT细胞IP-10表达量在培养 15 min内迅速下降,并且在 90 min的培养时间内保持在一个相当低的水平。虽然在培养至 90~ 120 min内, HaCaT细胞IP-10表达量有一定的升高,但是与空白组相比仍是受到抑制的。经统计学分析发现,与空白组相比,培养至 15、 30 min时, HaCaT细胞IP-10表达量显著降低(P< 0.05),而培养至 60、 90 min时,IP-10表达量极显著降低(P< 0.01),到培养 120 min时,IP-10表达量稍有回升,但仍显著低于空白组(P< 0.05)。以上结果表明,蓝莓叶总黄酮对IP-10基因的表达具有抑制作用。
炎症因子( IL-6、 IL-8)和趋化因子( CXCR-1和 CXCR-2、 IP-10)是反映炎症反应的重要指标,在机体抗感染、抑病毒过程中起重要作用 [15-16]。本实验将蓝莓叶总黄酮与 HaCaT细胞共培养特定时间后,利用 RT-qPCR检测相关炎症因子和趋化因子的表达情况。结果表明,蓝莓叶总黄 酮对IL-6 和CXCR-2 基因的表达具有正向调节的作用,二者的表达量显著增加。其中,IL-6通过影响细胞内花生四烯酸的代谢而起到抗炎作用,CXCR-2通过与配体IL-8结合,启动中性粒细胞趋化反应,募集中性粒细胞,引起炎症反应,对入侵的病原体发挥吞噬杀伤和清除作用。IL-8 和IP-10 基因的表达受到了明显抑制,表达量大幅度降低,进而抑制了炎症的发生和发展。 IP-10及其受体 CXCR-3具有强大的招募中性粒细胞能力,可促进多种细胞因子的分泌,作为趋化因子介导 Thl型炎症反应 [17-18],蓝莓叶总黄酮的添加抑制了 HaCaT细胞中 IP-10的表达,从而降低了炎症反应发生的概率。而蓝莓叶总黄酮对趋化因子CXCR-1的表达则呈现出复杂型调节的作用,表现为间歇性地促进和抑制,因此,蓝莓叶总黄酮具体的抗炎作用效果还有待研究。
综合来看,在炎症反应中有诸多的炎症因子和趋化因子参与到其中,这些因子之间相互作用,联系紧密,网络系统复杂多样,所以其表达量的变化必然会相互影响 [19-21]。整体来看,蓝莓叶总黄酮的添加会促进抗炎因子的表达,抑制促炎因子的表达,从而发挥其抗炎作用。但是蓝莓叶总黄酮的具体抗炎机理到目前为止还不是很清楚,有待进一步研究。本实验利用 RT-qPCR法对蓝莓叶总黄酮的抗炎功效进行了初步评价,为保健品或药食同源物质的体外抗炎评价提供了新思路。
参考文献:
[1] GERMOLEC D R, YOSHIDA T, GAIDO K, et al. Arsenic induces overexpression of growth factors in human keratinocytes[J]. Toxicology and Applied Pharmacology, 1996, 141(1)∶ 308-318.
[2] WEISFELNER M E, GOTTLIEB A B. The role of apoptosis in human epidermal keratinocytes[J]. Journal of Drugs in Dermatology, 2003, 2(4)∶ 385-391.
[3] 魏晶, 李伟, 邵万平, 等. CXCR1/CXCR2拮抗剂G31P抗中性粒细胞介导的炎症作用研究[J]. 中华微生物学和免疫学杂志, 2010(5)∶483-486.
[4] GRAS D, TIERS L, VACHIER I, et al. Regulation of CXCR/IL-8 in human airway epithelial cells[J]. International Archives of Allergy and Immunology, 2009, 152(2)∶ 140-150.
[5] HASHIZUME M, HIGUCHI Y, UCHIYAMA Y, et al. IL-6 plays an essential role in neutrophilia under inflammation[J]. Cytokine, 2011, 54(1)∶ 92-99.
[6] 蒋艳玲, 赵明, 梁功平, 等. 1,25-二羟维生素D 3对HaCaT细胞增殖活性及基因组DNA和增殖相关基因启动子甲基化水平的影响[J]. 中华皮肤科杂志, 2013, 46(12)∶ 885-888.
[7] 杨井, 陶娟, 李延, 等. 缺氧对HaCaT细胞HIF-1α, GLUT-1表达的影响及与细胞增殖的关系[J]. 中国皮肤性病学杂志, 2009, 23(10)∶621-623.
[8] 汤三妹, 杨娟, 周秋莲, 等. 猕猴桃RNA提取与RT-PCR[J]. 生物技术通报, 2006(5)∶ 67-71.
[9] 吴德全. IL-15调控HaCaT细胞增殖及分化的研究[D]. 合肥∶ 安徽医科大学, 2012∶ 14-36.
[10] TCHERAKIAN C, RIVAUD E, CATHERINOT E, et al. Pulmonary arterial hypertension related to HIV∶ is inflammation related to IL-6 the cornerstone?[J]. Revue de Pneumologie Clinique, 2011, 67(4)∶250-257.
[11] 许树长, 陈莹, 王锋, 等. IL-8及CXCR-1在反流性食管炎患者食管黏膜中的表达及意义[J]. 同济大学学报∶ 医学版, 2007, 28(5)∶ 51-54.
[12] FITZGERALD R C, ONWUEGBUSI B A, BAJAJ-ELLIOTT M, et al. Diversity in the oesophageal phenotypic response to gastrooesophageal reflux∶ immunological determinants[J]. Gut, 2002, 50(4)∶451-459.
[13] ISOMOTO H, WANG A, MIZUTA Y, et al. Elevated levels of chemokines in esophageal mucosa of patients with reflux esophagitis[J]. The American Journal of Gastroenterology, 2003, 98(3)∶ 551-556.
[14] AKCAY A, NGUYEN Q, EDELSTEIN C L. Mediators of inflammation in acute kidney injury[J]. Mediators of Inflammation, 2009. doi∶ 10.1155/2009/137072.
[15] 张如峰, 杨桂文, 安利国. 趋化因子及其受体在炎症中作用的研究进展[J]. 国际免疫学杂志, 2012, 35(3)∶ 191-196.
[16] 邢艳玲. IP-10对变应性接触性皮炎小鼠细胞因子表达影响的研究[D].天津∶ 天津医科大学, 2009∶ 15-31.
[17] FUENTE M, MIQUEL J. An update of the oxidation-inflammation theory of aging∶ the involvement of the immune system in oxiinflamm-aging [J]. Current Pharmaceutical Design, 2009, 15(26)∶3003-3026.
[18] 夏世金, 沈自尹, 董竞成. 老年大鼠下丘脑-垂体-肾上腺轴炎性衰老相关基因表达特征[J]. 老年医学与保健, 2009, 15(1)∶ 7-9.
[19] 郑梅竹. 罗布麻叶总黄酮抗抑郁作用及其机制研究[D]. 长春∶ 吉林大学, 2011∶ 40-106.
[20] SASAKI M, IKEDA H, SATO Y, et al. Proinflammatory cytokineinduced cellular senescence of biliary epithelial cells is mediated via oxidative stress and activation of ATM pathway∶ a culture study[J]. Free Radical Research, 2008, 42(7)∶ 625-632.
[21] BARTEK J, HODNY Z, LUKAS J. Cytokine loops driving senescence[J]. Nature Cell Biology, 2008, 10(8)∶ 887-889.
In vitro Anti-inflammatory Effect of Total Flavonoids from Blueberry Leaves
ZHAO Dan
1
, SU Ning
2
, YANG Li
2
, ZHENG Hongyan
2
, HUO Tong
1
, WANG Changtao
1,*
(1. School of Science, Beijing Technology and Business University, Beijing 100048, China; 2. Chinese Academy of Inspection and Quarantine, Beijing 100176, China)
Abstract: Purpose∶ To evaluate the in vitro anti-inflammatory potential of total flavonoids from blueberry leaves. Methods∶a co-culture system of HaCaT cell and flavonoids were constructed. To examine the effect of flavonoids on the expression of inflammatory cytokines including interleukin 6 (IL-6), interleukin-8 (IL-8), and interferon induced protein 10 (IP-10), realtime reverse transcription quantitative PCR (RT-qPCR) was used to detect the expression of the related genes after co-culture for different periods of time(0, 15, 30, 60, and 120 min). The anti-inflammatory mechanism of flavonoids from blueberry leaves was analyzed at the molecular level. Results∶ The total flavonoids from blueberry leaves significantly enhanced the expression of anti-inflammatory cytokines ( IL-6 and CXCR-2 ) ( P < 0.05) and significantly inhibited the expression of pro-inflammatory cytokines ( IL-8 , IP-10 ) ( P < 0.05) to achieve the anti-inflammatory activity. Chemotactic factor receptor CXCR-1 was regulated complicatedly. Conclusions∶ The total flavonoids from blueberry leaves could decrease inflammation by regulating the function of several gene loci via several steps.
Key words: total flavonoids from blueberry leaves; inflammatory cytokines; real-time reverse transcription PCR; anti-inflammatory
中图分类号: R966
文献标志码: A 文章编号:1002-6630(2015)17-0231-05
文章编号:1002-6630(2015)17-0231-05
doi:10.7506/spkx1002-6630-201517043
收稿日期:2014-11-05
基金项目:国家质检总局质检公益性行业科研专项(201310132;201410019)
作者简介:赵丹(1988—),女,助理实验师,硕士,研究方向为生物技术。E-mail:zhaodanustb@126.com
*通信作者:王昌涛(1975—),男,教授,博士,研究方向为植物功效成分开发应用。E-mail:wangct@th.btbu.edu.cn