多不饱和脂肪酸对大脑功能影响研究进展
刘志国,王丽梅,王华林,刘烈炬
*
(武汉轻工大学生物与制药工程学院,湖北 武汉 430023)
摘 要:多不饱和脂肪酸(polysaturated fatty acid,PUFA)是一类重要的脂肪酸族营养素,广泛参与细胞代谢和细胞膜脂的构成。在脑组织中,PUFA含量丰富,对促进大脑发育、增强学习记忆能力有重要作用,因而广受关注。本文综述PUFA的膳食来源及其与脑组织中磷脂构成的关系,以及PUFA参与大脑发育、影响大脑功能的作用机制,为确定膳食中ω-6/ω-3 PUFA的合理配比,科学添加源于深海鱼油和水藻的二十碳五烯酸(eicosapentaenoic acid,EPA)、二十二碳六烯酸(docosahexaenoic acid,DHA)或相关替代品,改善我国居民膳食中ω-3 PUFA的摄入不足,维持大脑健康功能提供理论依据。
关键词:多不饱和脂肪酸;磷脂;大脑发育;大脑功能
多不饱和脂肪酸是油脂中的一类重要的营养成分,其含量和种类决定了油脂的营养特性,对维护人类健康有重要作用,因而倍受人们的关注。大量研究证实,多不饱和脂肪酸(polysaturated fatty acid,PUFA)对维护细胞膜正常结构功能、参与细胞代谢调控、维持机体正常功能具有重要作用。其对神经系统的影响更是营养学与神经生物学共同关注的焦点。本文结合本课题组的研究
[1-4]与有关文献,针对PUFA对大脑功能的调节作用及机理做一综述,希望对引导合理健康饮食及油脂营养有所帮助。
1 PUFA的膳食来源与结构平衡
根据双键的位置不同,PUFA可分为ω-3和ω-6两种类型,又根据脂肪链的长度,超过18碳的被称为长链多不饱和脂肪酸(long-chain polyunsaturated fatty acid,LC-PUFA)。重要的ω-3 PUFA有α-亚麻酸(α-linolenic acid,ALA)、二十碳五烯酸(eicosapentaenoic acid,EPA)和二十二碳六烯酸(docosahexaenoic acid,DHA),ω-6 PUFA有亚油酸(linoleic acid,LA)、花生四烯酸(arachidonic acid,AA)等(表1)。近几十年,伴随着工业的快速发展和人们生活方式的改变,我国居民的膳食PUFA构成发生了很大变化。人们日常广泛使用含有高比例ω-6 PUFA,而缺乏ω-3 PUFA的大豆油、玉米油、花生油和葵花油等植物油(表2),导致食物中ω-6 PUFA含量明显增加,ω-3 PUFA含量相对降低
[5-6]。另外,肉类食品和奶类制品消费量的增加也是导致ω-6 PUFA摄入量增加的另一个重要原因。而ω-3 LC-PUFA,例如DHA和EPA的摄入则主要依赖于鱼类食品,特别是来自深海的鳕鱼、沙丁鱼和鲑鱼等鱼类产品
[7],少量来自植物源ALA转化生成的DHA、EPA,但转化率极低,通常约2%~10%,甚至更低
[8],因而导致我国居民ω-3 PUFA摄取普遍不足。而在美国,每个成年人平均的EPA和DHA摄入量大约也只有100 mg/d
[9]。
表1 常见的不饱和脂肪酸种类
Table 1 Common unsaturated fatty acids

注:MUFA. 单不饱和脂肪酸(monounsaturated fatty acid);DPA. 二十二碳五烯酸(docosapentaenoic acid)。
MUFAω-6 PUFAω-3 PUFA棕榈油酸(cis-C
16∶1
ω-7)亚油酸(cis-C
18∶2
ω-6)α-亚麻酸(cis-C
18∶3
ω-3)油酸(cis-C
18∶1
ω-9)γ-亚麻酸(cis-C
18∶3
ω-6)EPA(cis-C
20∶5
ω-3)反油酸(trans-C
18∶1
ω-9)花生四烯酸(cis-C
20∶4
ω-6)DPA(cis-C
22∶5
ω-3)芥子酸(cis-C
22∶1
ω-9)DPA(cis-C
22∶5
ω-6)DHA(cis-C
22∶6
ω-3)神经酸(cis-C
24∶1
ω-9)
表2 主要食用植物油中不饱和脂肪酸的构成
Table 2 Unsaturated fatty acid composition of edible vegetables oils %

序号油脂名称ω-3 PUFAω-6 PUFAω-9 PUFA序号油脂名称ω-3 PUFAω-6 PUFAω-9 PUFA 1大豆油4~650~6020~25 11米糠油1~230~4040~45 2菜籽油5~810~2040~6012棉籽油0~155~6015~20 3花生油0~330~4030~4013南瓜籽油055~6015~20 4橄揽油0~310~1568~7514紫苏籽油51~6312~2013~22 5玉米胚芽油1~360~6520~2515亚麻籽油45~6110~1515~20 6葵花籽油0~165~7018~2216核桃油6~855~6515~20 7茶籽油0~35~1075~8617鲸鱼油(长须鲸内部脂)12.92.460.9 8红花油0~175~858~1018步鱼油27.42.323.9 9棕榈油0~115~2035~4519飞鱼油27.57.729.2 10芝麻油1~345~5035~4020鳕鱼肝油25.12.544.9
饮食结构改变所致的ω-6/ω-3摄入比例失衡已成为现代社会人们重要的饮食特征。近几十年来,西方国家居民ω-6 LC-PUFA的摄入量(主要是AA)增加了1 倍多,ω-6/ω-3摄入比例随之升高。在法国,饮食中的LA/ALA比例增加了4 倍,达到10∶1~15∶1,远高于膳食营养指南推荐的小于5∶1的要求
[10-11]。LA和ALA是LC-PUFA合成的前体,由于人和哺乳动物自身不能合成LA和ALA,食物中的LA/ALA比例决定了人体ω-6/ω-3 PUFA的比例。虽然ω-6与ω-3 PUFA在体内不能相互转化,然而ω-6和ω-3 LC-PUFA的体内合成却使用共同的延长酶和去饱和酶,使ω-6和ω-3 LC-PUFA的合成具有竞争关系(图1)。研究发现,饮食中限制ω-6 PUFA的摄入量,能使ALA至DHA转换率增加25%
[12]。值得注意的是,由于食物中ω-3 LC-PUFA相对缺乏,EPA和DHA合成效率极低,饮食中高比例的ω-6 PUFA可对ω-3 LC-PUFA的合成造成显著的竞争性抑制效应,导致体内ω-3 LC-PUFA含量进一步降低。饮食中ω-3 PUFA摄入量过低以及ω-6/ω-3比例过高可影响血液、红细胞、乳汁、脂肪组织以及大脑中ω-6和ω-3 PUFA的含量
[13],增加多种疾病发生的风险。
研究显示,较高的ω-6/ω-3 PUFA摄入比例或ω-3 PUFA缺乏可导致代谢紊乱,是诱发神经系统疾病、心血管疾病、癌症等慢性疾病的重要原因之一
[14-17]。流行病学研究显示,饮食中ω-6/ω-3 PUFA比例过高与智力发育迟缓、老年痴呆等疾病的高发生率密切相关
[18]。此外,高ω-6/ω-3 PUFA比例可促进人类前列腺肿瘤细胞的生长
[19],并增加乳腺癌和胰腺癌发病风险
[20],并对精子有显著损伤作用,降低精子的数量和质量
[21-22]。因此,在现代饮食中摄入足够量的ω-3 PUFA,并保持ω-6/ω-3 PUFA摄入量的平衡显得尤为重要。

图1 长链不饱和脂肪酸的合成途径
[14]
Fig.1 Synthesis pathway of long-chain polyunsaturated fatty acids
[14]
2 脑组织PUFA的构成
2.1 脑组织PUFA的分布特点
ω-3和ω-6 PUFA占脑组织脂肪酸总量的30%~35%,并主要以与磷脂结合的形式存在。在哺乳动物中,磷脂占脑组织干质量的50%~60%。PUFA在脑组织具有不对称分布的特点,表现为不同种类的PUFA在脑组织中含量不同:高含量的DHA和AA,低含量的EPA、ALA和LA
[23](图2)。PUFA构成的磷脂在细胞膜脂质双层结构中也呈不对称分布。在哺乳动物的细胞膜上,磷脂酰胆碱(phosphatidylcholine,PC)多集中在脂质双分子层的外侧小叶,而磷脂酰丝氨酸(phosphatidylserine,PS)、磷脂酰乙醇胺(phosphatidylethanolamine,PE)和磷脂酰肌醇(phosphatidylinositol,PI)更集中于内侧小叶
[24]。虽然对这些PUFA不对称分布的生理意义目前尚缺乏充分研究,但很显然,这与细胞膜脂质双层结构的流动性、可塑性以及脂筏的形成有关,同时也可能影响到脂质双层结构中蛋白质(例如酶、受体和通道蛋白等)的功能
[25]。
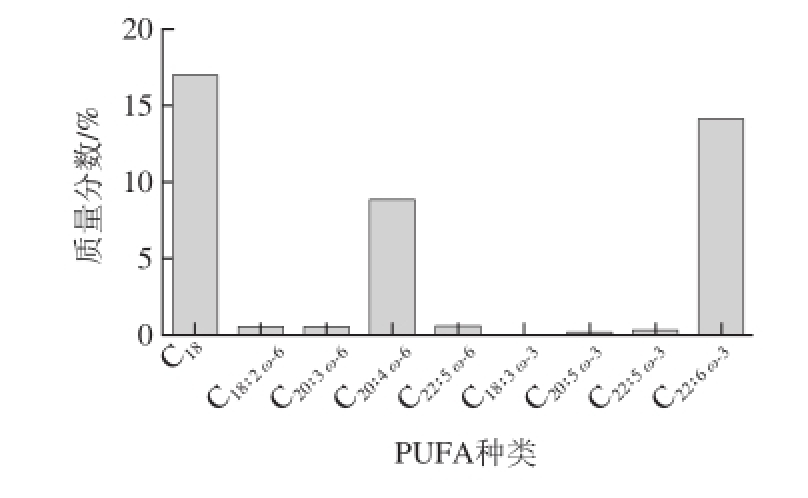
图2 脑磷脂中不同脂肪酸的含量
Fig.2 Contents of different fatty acids in brain phospholipids
引自参考文献[14],并略作修改。
为了保持细胞膜PUFA水平的相对稳定,大脑需要不断从血液摄取来自饮食和肝脏合成的脂肪酸。过去对PUFA通过血脑屏障的方式一直存有争议。曾经有观点认为PUFA通过血脑屏障需要脂蛋白或血浆蛋白等转运机制的参与。但最近的研究采用原位脑灌注技术发现EPA和DHA可以脂溶性的方式迅速通过血脑屏障
[26-27]。然而实验发现大脑磷脂中DHA的含量是EPA的300 倍
[28]。如何解释脂肪酸在大脑中的不均匀分布?最近的研究发现:在给成年大鼠饲喂富含DHA和ALA的饲料后,虽然ALA可以迅速从血浆扩散到大鼠的大脑,但大部分ALA通过β-氧化而降解,只剩下10%用于磷脂的合成,2%用于合成甘油三酯,仅≤0.2%用以合成新的DHA
[29]。其他研究发现,86%以上从血浆摄取的LA在大脑中被β-氧化,只有<1%的部分用于合成AA。给大鼠侧脑室注入放射性元素标记的EPA,4 d之后,发现只有9%完整的EPA,剩余的均被β-氧化或去酯化降解
[28]。这些研究表明,脂肪酸在脑中的不均匀分布可能与其差异化的降解和酯化过程有关。
在大鼠饲料缺乏DHA的情况下,肝脏ALA转化为DHA的速率加快,以维持大脑正常的需要。因此在肝功能损伤的情况下或老年性肝脏功能衰退时,需要从膳食中直接摄取更多的DHA,以维持大脑中DHA的水平。实验发现神经细胞膜ω-3 PUFA含量会随着年龄增长和神经变性疾病而减少
[30]。尽管神经细胞质膜的成分能在一定范围可通过自身的调节机制保持相对稳定,但在很大程度上仍受到饮食的影响。例如在为小鼠补充EPA 6~8 周后,可以显著增加其神经细胞膜EPA和代谢物DPA含量
[31]。长期补充DHA可增加大脑的DHA含量,相应降低大脑ω-6 PUFA的含量。相反,在饮食中长期缺乏ω-3 PUFA,将耗尽帕金森模型小鼠大脑中的DHA,伴随大脑ω-6 PUFA的含量相应的增加
[32]。ω-6/ω-3 PUFA比例严重失调可造成大脑功能损伤的潜在风险
[33]。
2.2 PUFA在膜磷脂中的特异性分布
磷脂是构成神经和胶质细胞膜脂质双层结构的主体。进入脑组织的PUFA(包括DHA和AA)在酰基辅酶A合成酶的作用下转化为酰基辅酶A,然后通过酰基辅酶A转移酶形成甘油磷脂。
在人类的大脑灰质、白质和髓鞘中,不同的甘油磷脂所占干质量的百分比不同。整体而言,胆碱甘油磷脂(choline glycerophosphates,CGP)和乙醇胺甘油磷脂(ethanolamine glycerophosphates,EGP)含量较高,而丝氨酸甘油磷脂(serine glycerophosphates,SGP)含量较低。有实验研究了不同的脂肪酸在甘油磷脂中的分布,发现最大比例的PUFA(40%~60%)存在于大脑灰质的EGP和SGP中,只微量存在于CGP中;不同于甘油磷酸酯,棕榈酸在CGP中含量最高,而在SGP中含量极低。在鞘磷脂中,PUFA的含量最低(特别是在CGP和SGP中),而油酸的含量最高
[34]。研究发现脂肪酸在磷脂中的特异性分布具有重要的生理意义。例如SGP中DHA的含量与神经细胞的生存能力密切相关,可能成为判断神经细胞生存能力的一个指标。当大脑DHA含量不足时,DPA(ω-6)即可取代PS中的DHA,造成神经细胞的损伤
[35-36]。此外,尽管CGP中DHA的含量很低,但随着年龄的增长,大脑灰质CGP中的DHA的含量却迅速增加,与老年大脑的衰老过程密切相关
[37]。目前已有许多研究对饮食中EPA和DHA与血浆以及红细胞胞膜中EPA和DHA的关联性进行了探讨
[38],然而迄今还没有任何直接的数据揭示膳食中脂肪酸与大脑中脂肪酸的相关性
[14]。脂肪酸在磷脂中的特异性分布及其生理意义将是今后研究探讨的重要方向。
磷脂的合成需要多种前体,包括尿苷、脂肪酸和胆碱(图3)。实验发现,沙鼠单独摄入尿嘧啶不能增加磷脂的含量(PC、PE、PS和PI)。沙鼠单独摄入DHA在某种程度上可以增加一些磷脂(PE、PS和PI)的水平,但不能增加PC水平。然而,沙鼠尿嘧啶和DHA联用,可以增加28%的PC含量。单独摄入DHA仅可增加26%的PE含量,结合尿苷使用则可增加59%的含量。单独使用DHA仅增加75%的PS含量,结合尿苷使用可达到160%。单独使用DHA仅增加了29%的PI含量,结合尿苷的使用可增加到100%
[39]。上述结果表明,DHA和尿苷对增加磷脂的合成具有协同作用,同时也提示了两者联合干预的可行性和必要性。值得注意的是,在大脑脂肪酸的积累过程中存在一种层级结构,例如进入大脑的DHA总是优先结合在大脑灰质的EGP和SGP中,而不是CGP中
[34]。DHA对不同的磷脂合成也具有优先的层级结构,即最高的层级是PS、其后是PI、然后是PE和PC。采用神经嗜铬细胞瘤细胞培养方法,发现DHA可被细胞摄取,并可激活酰基辅酶A合成酶,合成富含DHA的磷脂
[40]。磷脂合成的增加可促进神经突起的生长,增加突触和树突棘的数量,促进大脑的学习记忆功能
[41]。
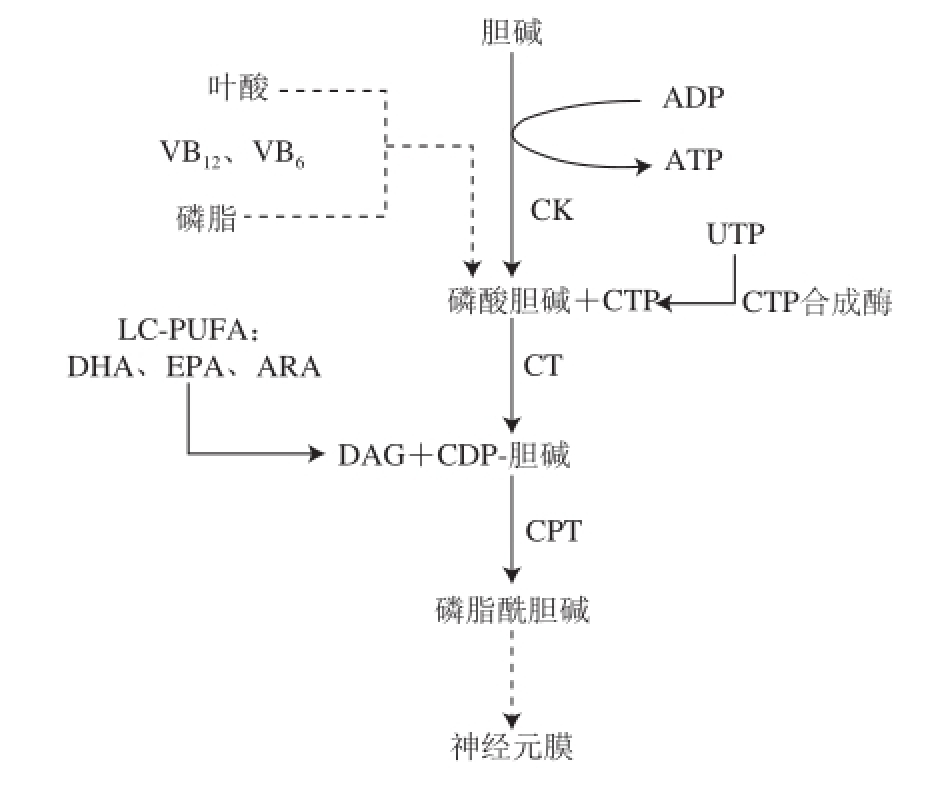
图3 神经细胞膜的形成
Fig.3 Formation of the nerve cell membrane
引自参考文献[34]并略作修改。从LC-PUFA到神经元质膜的形成,被称为肯尼迪途径或肯尼迪循环。ADP. 二磷酸腺苷(adenosine diphosphate);ARA. 花生四烯酸(arachidonic acid);ATP. 三磷酸腺苷(adenosine triphosphate);CDP-choline. 二磷酸胞苷-胆碱(cytidine diphosphate choline);CK. 胆碱激酶(choline kinase);CPT. 甘油二酯磷酸转移酶(1,2-diacylglycerol choline phosphotransferase);CT. 三磷酸胞嘧啶转移酶(cytidine triphosphate-phosphocholine cytidyl transferase);CTP. 三磷酸胞嘧啶(cytidine triphosphate);DAG. 甘油二酯(diacylglycerol);UTP. 三磷酸尿苷(uridine triphosphate)。
总之PUFA和磷脂的关系越来越受到关注。首先磷脂是PUFA的重要载体,在磷脂中PUFA呈特异性分布,具有重要的生理意义。另外饮食中的PUFA和尿苷不仅可以协同增加各自在大脑中的含量,同时对促进大脑磷脂的合成具有协同作用,提示了一种PUFA联合干预的新途径。
3 PUFA对大脑功能的影响及机制
除脂肪组织外,大脑是PUFA(特别是DHA)含量的最高器官。PUFA对维持大脑正常的结构和功能起着重要作用。PUFA在婴幼儿期可以促进大脑的生长发育;在老年期可延缓大脑的衰老。虽然对PUFA的作用目前仍有争论
[42-43],但研究者普遍认为ω-3 PUFA的缺乏有碍于大脑正常功能;相反,适当地补充ω-3 PUFA则有益于大脑正常的结构和功能的维护
[44]。越来越多证据表明:饮食中ω-3 PUFA的摄入量还与多种神经和精神性疾病有关,包括婴幼儿时期神经发育障碍,如多动症等
[45]以及中老年期的阿尔茨海默病和帕金森病等
[46-47]。目前全球精神和神经系统疾病的发病率已经超过了心血管疾病和癌症的发病率,迫切需要有一种新的、安全有效的防治方法,PUFA提供了这样一种潜在的可能性
[48]。我国目前已进入了老年化的社会,伴随而来的老年人学习记忆或认知能力的衰退已日益成为严重的社会问题
[49]。因此,PUFA增强大脑学习记忆的作用以及作用机理是目前营养生物学和食品科学探讨的重要课题
[50]。
3.1 PUFA与大脑发育
人类大脑快速生长期开始于妊娠晚期,在此期间胎儿大脑的DHA开始大量集聚。快速生长期一直延续到出生后的第24个月,出生后的婴儿期DHA仍在快速聚集,但相比妊娠晚期,聚集速率开始有所下降
[51]。LC-PUFA是维持大脑和视觉系统生长发育的必需营养素
[52]。胎盘中脂肪酸的含量取决于母亲血液中脂肪酸的含量
[53];出生后婴儿的PUFA来源于母乳,支持婴儿期大脑的快速生长发育。虽然大脑可以合成少量的LC-PUFA,但AA和DHA主要来源于血液,肝脏是人体合成AA和DHA的主要器官。大多数AA和DHA进入大脑是在胎儿发育晚期和产后早期,这是补充ω-6和ω-3 PUFA的最关键时期。针对人和动物的研究均表明围产期母体饮食中ω-3 PUFA的缺乏可导致胎儿大脑DHA含量的下降。在怀孕期间雌性大鼠ω-3 PUFA摄入量的不足,可导致幼鼠大脑DHA的含量与正常大鼠相比下降50%
[54]。大量研究表明婴儿期是摄取ω-3 PUFA的重要时期,尤其是DHA。研究发现,与母乳喂养的婴儿相比,缺乏ω-3 PUFA的摄入可使一些婴儿大脑DHA的含量下降35%
[55],因此在婴儿配方奶粉中对PUFA的补充应引起人们的高度重视
[56]。
3.2 PUFA与大脑功能
对成年人而言,大脑的DHA含量可直接影响细胞膜的特性、突触的生成和可塑性,从而影响大脑学习记忆的功能
[44,57]。在磷脂酶A
2(phospholipase A
2,PLA
2)的作用下,大脑质膜,包括神经和神经胶质细胞的细胞膜不断释放非酯化的AA和DHA。这些被释放的PUFA有以下主要的代谢途径:1)转化为重要的生物活性分子,如花生四烯酸、白细胞三烯、凝血恶烷和前列腺素等;2)通过β-氧化,进入能量代谢途径或重新酯化形成新的膜磷脂;3)扩散到血液,在大脑和血浆之间进行交换。实验表明大鼠每天PUFA的交换率大约为5%~8%。人类AA交换率为18 mg/d,DHA交换率为4~5 mg/d
[58-59]。可见,为了维持大脑正常的结构和功能,每天必须要消耗一定数量的DHA,被消耗的DHA需要由饮食中的ω-3 PUFA来补充。在饲料中限制ω-3 PUFA摄入量,4 个月后成年大鼠大脑中DHA的含量低于正常大脑的37%
[60]。因此根据我国居民目前的饮食习惯,饮食中ω-3 PUFA的缺乏可能会导致一些个体大脑DHA含量的不足,从而影响大脑的结构和功能。当DHA缺乏时,大脑可启动有效的反馈机制,减少DHA的消耗量。实验发现,当饲料中ω-3 PUFA不足时,DHA在成年大鼠的脑和血浆之间的交换率可延长至4 个月,交换率低于正常对照组的4 倍
[60]。大脑DHA的缺乏可触发一系列的信号级联反应,并导致DHA和AA之间的代谢失衡。AA及其代谢产物过剩可促进大脑的炎性反应。实验证明膳食中ω-3 PUFA的摄入量可以影响体内PLA
2/COX axis和脂质信号转导系统的活动
[61]。在大脑衰老和神经系统炎性反应过程中,神经细胞、小胶质细胞和星形胶质细胞膜上的PLA
2/COX axis是维持大脑内环境稳定的重要因素
[62]。因此在成年期饮食中ω-3 PUFA摄入量不足不仅可导致大脑DHA的含量逐渐减少,影响大脑的功能;同时也可能为老年期大脑慢性炎症的发生埋下隐患
[63]。
随着年龄的增长,大脑逐渐衰老,脑内的DHA含量开始下降。实验显示老年大鼠细胞膜的DHA含量明显低于年轻大鼠
[64]。DHA在不同膜磷脂中的含量降幅在5%~20%之间
[65]。膳食中ω-3 PUFA的不足会加速大脑的老年化过程,大脑的老年化过程又进一步导致大脑DHA的流失。最新的一些研究发现,由年龄导致的大脑DHA的流失可能与一些酶类活性下降有关,例如长链酰基辅酶A合成酶,该酶可选择性地催化DHA与大脑膜磷脂结合
[66]。实验证明大脑DHA缺乏不仅是衰老导致大脑学习记忆功能下降重要原因;同时与多种和学习记忆相关的老年性疾病有关
[46-47]。
总之,PUFA在大脑的生长发育、功能成熟和衰老进程中起着重要的作用。在婴幼儿期,PUFA促进大脑的生长发育;在成年期,PUFA维持大脑的正常功能;在老年期,PUFA可延缓大脑的衰老。
3.3 PUFA影响大脑功能的可能机制
PUFA对大脑的作用,除了以磷脂为载体,构成细胞膜脂质双层结构,影响细胞膜的流动性和可塑性以外,还可通过调节局部游离脂肪酸以及磷脂信号分子的浓度而发挥生理效应。如前文所述,大脑中的PUFA主要来源于食物,体内最大的PUFA合成器官是肝脏。但局部游离脂肪酸和脂质信号分子的浓度除了取决于整体的PUFA浓度外,还依赖于局部质膜的释放。细胞膜的构成和局部游离脂酸的浓度是影响细胞信号转导系统的两个重要因素。人们通常把细胞质膜形容为动态游离脂肪酸和脂质信号分子的储库
[25]。细胞膜脂质的构成和磷脂中脂肪酸的特性可直接影响质膜结构和脂质信号转导系统
[26]。脂肪酸的长度和饱和度可直接影响脂质双层的侧向扩散运动和蛋白质在脂质双层结构中的流动性
[67]。不饱和脂肪酸的双键结构在质膜的流动性和可塑性中扮演着重要角色
[68]。在神经递质、神经营养因子和细胞因子的作用下,细胞膜可通过酶解反应释放出多种脂质信号分子,提高大脑脂质的可用度,如磷酯酰肌醇二磷酸、脂肪酸酯辅酶A、溶血磷脂酸和游离的脂肪酸等可改善细胞的功能。长期不良饮食、疾病、生长发育障碍和衰老均可引起神经细胞膜结构的改变和脂质可用度的降低。
细胞质膜释放游离脂肪酸的过程可能与炎性作用有关,本文仅以AA为例来说明细胞质膜PUFA的释放过程和对PUFA可用度调节。激活某些G蛋白偶联代谢型受体,可引起细胞膜磷脂的酶解反应,释放出游离脂肪酸。参与这些酶解反应的有3 个重要的磷脂酶,即磷脂酶A
2(PLA
2)、磷脂酶C(phospholipase C,PLC)和磷脂酶D(phospholipase D,PLD)。上述3 种磷脂酶分别作用于磷脂中的不同基团,产生不同的酶解产物。在哺乳动物细胞中包含多种PLA
2的同工酶。PLA
2可直接催化磷脂质sn-2脂酰键的水解,产生AA、DHA和溶血磷脂
[69]。PLC和PLD的激活不能直接产生AA,但可以通过AA的前体——二脂酰甘油或磷脂酸释放AA。释放出的AA可能成为逆行信使(retrograde messenger)
[70],或进一步通过各种代谢酶(如环氧酶、脂氧合酶和加双氧酶等)生成多种类花生酸物质,如前列腺素、白细胞三烯、凝血恶烷等。类花生酸在机体炎性反应中起着重要的作用。不同的是,类花生酸主要产生的是促炎因子,而DHA主要产生抗炎因子
[71-72]。
因此,大脑中游离脂肪酸的浓度,特别局部游离脂肪酸的浓度,不仅仅取决于食物中ω-3 PUFA的含量,还取决于细胞质膜对游离脂肪酸的释放。PUFA对调节大脑的功能具有重要作用。在此过程中,PLA
2、PLC和PLD是关键酶,是PUFA作用的靶点。
4 结 语
综上所述,PUFA与人体健康和大脑功能有着密切的关系。大量摄取富含ω-3 PUFA的饮食既可以降低缺血性心脏病的风险,又有助于大脑的健康
[73-74]。目前我国居民的膳食中并不缺乏ω-6 PUFA,而ω-3 PUFA的摄入量相对不足。与西方国家相比,我国居民的鱼类食品,特别是海洋鱼类食品摄入量较少,表现为ω-3 LC-PUFA摄入量更加匮乏。这些膳食特点应该对油脂工业和食品科学研究领域有所启示。世界卫生组织下设的油脂委员会和中国营养学会提出了食用油中饱和脂肪酸、单不饱和脂肪酸、多不饱和脂肪酸的组成比例应为1∶1∶1的建议
[75]。此外,最新的一系列研究发现食用油中ω-3 LC-PUFA的含量以及ω-6/ω-3 PUFA的比例对食用油的营养价值具有同样重要的意义。对我国的消费者和生产者而言,重视食用油中添加来源于深海鱼油和水藻的EPA和DHA,以及寻找和挖掘EPA的DHA的替代品对维护健康具有重要意义。
参考文献:
[1] PAN J P, ZHANG H Q, WEI W, et al. Some subtypes of endocannabinoid/endovanilloid receptors mediate docosahexaenoic acid-induced enhanced spatial memory in rats[J]. Brain Research, 2011, 1412: 18-27.
[2] 王丽梅, 叶诚, 郭艳芬, 等. DHA油与其他4种油脂对大鼠空间学习记忆、脂质过氧化及其血脂水平的影响[J]. 中国油脂, 2013, 38(3): 34-38.
[3] 马琳, 王丽梅, 樊永波, 等. DHA提高大鼠学习记忆的分子机制研究[J]. 食品科学, 2013, 34(17): 246-249. doi: 10.7506/spkx1002-6630-201317052.
[4] 郭艳芬, 王丽梅, 肖娜, 等. 发育期补充DHA对大鼠空间学习记忆能力的影响[J]. 中国油脂, 2011, 36(9): 43-47.
[5] LEAF A, WEBER P C. A new era for science in nutrition[J]. American Journal of Clinical Nutrition, 1987, 45(Suppl 5): 1048-1053.
[6] BILTON R. Averting comfortable lifestyle crises[J]. Science Progress, 2013, 96(4): 319-368.
[7] STONE N J. Fish consumption, fish oil, lipids, and coronary heart disease[J]. Circulation, 1996, 94(9): 2337-2340.
[8] KELLY L, GREHAN B, CHIESA A D, et al. The polyunsaturated fatty acids, EPA and DPA exert a protective effect in the hippocampus of the aged rat[J]. Neurobiology of Aging, 2011, 32(12): 2318.e1-2318.e15. doi: 10.1016/j.neurobiolaging.2010.04.001.
[9] ERVIN R B, WRIGHT J D, WANG C Y, et al. Dietary intake of fats and fatty acids for the United States population: 1999-2000[J]. Advance Data, 2004, 348: 1-6.
[10] AILHAUD G, MASSIERA F, WEILL P, et al. Temporal changes in dietary fats: role of n-6 polyunsaturated fatty acids in excessive adipose tissue development and relationship to obesity[J]. Progress in Lipid Research, 2006, 45(3): 203-236.
[11] LEGRAND P, SCHMITT B, MOUROT J, et al. The consumption of food products from linseed-fed animals maintains erythrocyte omega-3 fatty acids in obese humans[J]. Lipids, 2010, 45(1): 11-19.
[12] BAZINET R P, DOUGLAS H, CUNNANE S C. Whole-body utilization of n-3 PUFA in n-6 PUFA-deficient rats[J]. Lipids, 2003, 38(2): 187-189.
[13] POTTALA J V, YAFFE K, ROBINSON J G, et al. Higher RBC EPA + DHA corresponds with larger total brain and hippocampal volumes: WHIMS-MRI study[J]. Neurology, 2014, 82(5): 435-442.
[14] LUCHTMAN D W, SONG C. Cognitive enhancement by omega-3 fatty acids from child-hood to old age: findings from animal and clinical studies[J]. Neuropharmacology, 2013, 64: 550-565.
[15] JANSSEN C I, KILIAAN A J. Long-chain polyunsaturated fatty acids (LCPUFA) from genesis to senescence: the influence of LCPUFA on neural development, aging, and neurodegeneration[J]. Progress in Lipid Research, 2014, 53: 1-17.
[16] MONTEIRO J, LESLIE M, MOGHADASIAN M H, et al. The role of n-6 and n-3 polyunsaturated fatty acids in the manifestation of the metabolic syndrome in cardiovascular disease and non-alcoholic fatty liver disease[J]. Food & Function, 2014, 5(3): 426-435.
[17] YANG B, REN X L, FU Y Q, et al. Ratio of n-3/n-6 PUFAs and risk of breast cancer: a meta-analysis of 274135 adult females from 11 independent prospective studies[J]. BMC Cancer, 2014, 14(2): 105. doi: 10.1186/1471-2407-14-105.
[18] DULLEMEIJER C, DURGA J, BROUWER I A, et al. n-3 fatty acid proportions in plasma and cognitive performance in older adults[J]. American Journal of Clinical Nutrition, 2007, 86(5): 1479-1485.
[19] ROSE D P, CONNOLLY J M. Omega-3 fatty acids as cancer chemopreventive agents[J]. Pharmacology & Therapeutics, 1999, 83(3): 217-244.
[20] FUNAHASHI H, SATAKE M, HASAN S, et al. Opposing Effects of n-6 and n-3 polyunsaturated fatty acids on pancreatic cancer growth[J]. Pancreas, 2008, 36(4): 353-362.
[21] FAIR S, DOYLE D N, DISKIN M G, et al. The effect of dietary n-3 polyunsaturated fatty acids supplementation of rams on semen quality and subsequent quality of liquid stored semen[J]. Theriogenology, 2014, 81(2): 210-219.
[22] YAN L, BAI X L, FANG Z F, et al. Effect of different dietary omega-3/ omega-6 fatty acid ratios on reproduction in male rats[J]. Lipids in Health and Disease, 2013, 12: 33. doi: 10.1186/1476-511X-12-33.
[23] PALSDOTTIR V, MANSSON J E, BLOMQVIST M, et al. Long-term effects of perinatal essential fatty acid deficiency on anxiety-related behavior in mice[J]. Behavioral Neuroscience, 2012, 126(2): 361-369. [24] DEVAUX P F. Protein involvement in transmembrane lipid asymmetry[J]. Annual Review of Biophysics and Biomolecular Structure, 1992, 21: 417-439.
[25] BOLAND L M, DRZEWIECKI M M. Polyunsaturated fatty acid modulation of voltage-gated ion channels[J]. Cell Biochemistry and Biophysics, 2008, 52(2): 59-84.
[26] OUELLET M, EMOND V, CHEN C T, et al. Diffusion of docosahexaenoic and eicosapentaenoic acids through the bloodbrain barrier: an in situ cerebral perfusion study [J]. Neurochemistry International, 2009, 55(7): 476-482.
[27] GREEN J T, LIU Z, BAZINET R P. Brain phospholipid arachidonic acid half-lives are not altered following 15 weeks of n-3 polyunsaturated fatty acid adequate or deprived diet[J]. Journal of Lipid Research, 2010, 51(3): 535-543.
[28] CHEN C T, LIU Z, BAZINET R P. Rapid de-esterification and loss of eicosapentaenoic acid from rat brain phospholipids: an intracerebroventricular study[J]. Journal of Neurochemistry, 2011, 116(3): 363-373.
[29] DEMAR J C, MA K, CHANG L, et al. α-Linolenic acid does not contribute appreciably to docosahexaenoic acid within brain phospholipids of adult rats fed a diet enriched in docosahexaenoic acid[J]. Journal of Neurochemistry, 2005, 94(4): 1063-1076.
[30] YEHUDA S, RABINOVITZ S, CARASSO R L, et al. The role of polyunsaturated fatty acids in restoring the aging neuronal membrane[J]. Neurobiology of Aging, 2002, 23(5): 843-853.
[31] LUCHTMAN D W, MENG Q, SONG C. Ethyl-eicosapentaenoate (E-EPA) attenuates motor impairments and inflammation in the MPTP-probenecid mouse model of Parkinson’s disease[J]. Behavioural Brain Research, 2012, 226(2): 386-396.
[32] BOUSQUET M, SAINT-PIERRE M, JULIEN C, et al. Beneficial effects of dietary omega-3 polyunsaturated fatty acid on toxin-induced neuronal degeneration in an animal model of Parkinson’s disease[J]. FASEB Journal, 2008, 22(4): 1213-1225.
[33] SIMOPOULOS A P. The importance of the ratio of omega-6/omega-3 essential fatty acids[J]. Biomedicine & Pharmacotherapy, 2002, 56(8): 365-379.
[34] PARLETTA N, MILTE C M, MEYER B J. Nutritional modulation of cognitive function and mental health[J]. Journal of Nutritional Biochemistry, 2013, 24(5): 725-743.
[35] KIM H Y, BIGELOW J, KEVALA J H. Substrate preference in phosphatidylserine biosynthesis for docosahexaenoic acid containing species[J]. Biochemistry, 2004, 43(4): 1030-1036.
[36] KIM H Y. Novel metabolism of docosahexaenoic acid in neural cells[J]. Journal of Biological Chemistry, 2007, 282(26): 18661-18665.
[37] SCHAEFER E J, BONGARD V, BEISER A S, et al. Plasma phosphatidylcholine docosahexaenoic acid content and risk of dementia and Alzheimer disease: the framingham heart study[J]. Archives of Neurology, 2006, 63(11): 1545-1550.
[38] KROGER E, VERREAULT R, CARMICHAEL P H, et al. Omega-3 fatty acids and risk of dementia: the Canadian study of health and aging[J]. American Journal of Clinical Nutrition, 2009, 90(1): 184-192.
[39] SAKAMOTO T, CANSEV M, WURTMAN R J. Oral supplementation with docosahexaenoic acid and uridine-5’-monophosphate increases dendritic spine density in adult gerbil hippocampus[J]. Brain Research, 2007, 1182: 50-59.
[40] MARSZALEK J R, KITIDIS C, DIRUSSO C C, et al. Long-chain acyl-CoA synthetase 6 preferentially promotes DHA metabolism[J]. Journal of Biological Chemistry, 2005, 280(11): 10817-10826.
[41] WURTMAN R J, ULUS I H, CANSEV M, et al. Synaptic proteins and phospholipids are increased in gerbil brain by administering uridine plus docosahexaenoic acid orally[J]. Brain Research, 2006, 1088(1): 83-92.
[42] GOULD J F, MAKRIDES M, COLOMBO J, et al. Randomized controlled trial of maternal omega-3 long-chain PUFA supplementation during pregnancy and early childhood development of attention, working memory, and inhibitory control[J]. American Journal of Clinical Nutrition, 2014, 99(4): 851-859.
[43] DANTHIIR V, HOSKING D, BURNS N R, et al. Cognitive performance in older adults is inversely associated with fish consumption but not erythrocyte membrane n-3 fatty acids[J]. Journal of Nutrition, 2014, 144(3): 311-320.
[44] STONEHOUSE W, CONLON C A, PODD J, et al. DHA supplementation improved both memory and reaction time in healthy young adults: a randomized controlled trial[J]. American Journal of Clinical Nutrition, 2013, 97(5): 1134-1143.
[45] KURATKO C N, BARRETT E C, NELSON E B, et al. The relationship of docosahexaenoic acid (DHA) with learning and behavior in healthy children: a review[J]. Nutrients, 2013, 5(7): 2777-2810.
[46] DACKS P A, SHINEMAN D W, FILLIT H M. Current evidence for the clinical use of long-chain polyunsaturated n-3 fatty acids to prevent age-related cognitive decline and Alzheimer’s disease[J]. Journal of Nutrition, Health & Aging, 2013, 17(3): 240-251.
[47] CEDERHOLM T, SALEM N, PALMBLAD J. Omega-3 fatty acids in the prevention of cognitive decline in humans[J]. Advances in Nutrition, 2013, 4(6): 672-676.
[48] COLLINS P Y, PATEL V, JOESTL S S, et al. Grand challenges in global mental health[J]. Nature, 2011, 475: 27-30.
[49] SIERRA S, ALFARO J M, SANCHEZ S, et al. Administration of docosahexaenoic acid before birth and until aging decreases kainateinduced seizures in adult zebrafish[J]. Brain Research Bulletin, 2012, 88(5): 467-470.
[50] CRUPI R, MARINO A, CUZZOCREA S. n-3 Fatty acids: role in neurogenesis and neuroplasticity[J]. Current Medicinal Chemistry, 2013, 20(24): 2953-2963.
[51] DAGAI L, PERI-NAOR R, BIRK R Z. Docosahexaenoic acid significantly stimulates immediate early response genes and neurite outgrowth[J]. Neurochemical Research, 2009, 34(5): 867-875.
[52] KOLETZKO B, LIEN E, AGOSTONI C, et al. The roles of long-chain polyunsaturated fatty acids in pregnancy, lactation and infancy: review of current knowledge and consensus recommendations[J]. Journal of Perinatal Medicine, 2008, 36(1): 5-14.
[53] LO A, SIENNA J, MAMAK E, et al. The effects of maternal supplementation of polyunsaturated fatty acids on visual, neurobehavioural, and developmental outcomes of the child: a systematic review of the randomized trials[J]. Obstetrics and Gynecology International, 2012. doi: 10.1155/2012/591531.
[54] CHAMPEIL-POTOKAR G, CHAUMONTET C, GUESNET P, et al. Docosahexaenoic acid (22:6 n-3) enrichment of membrane phospholipids increases gap junction coupling capacity in cultured astrocytes[J]. European Journal of Neuroscience, 2006, 24(11): 3084-3090.
[55] FARQUHARSON J, JAMIESON E C, ABBASI K A, et al. Effect of diet on the fatty acid composition of the major phospholipids of infant cerebral cortex[J]. Archives of Disease in Childhood, 1995, 72(3): 198-203.
[56] TAI E K, WANG X B, CHEN Z Y. An update on adding docosahexaenoic acid (DHA) and arachidonic acid (AA) to baby formula[J]. Food & Function, 2013, 4(12): 1767-1775.
[57] HENNEBELLE M, CHAMPEIL-POTOKAR G, LAVIALLE M, et al. Omega-3 polyunsaturated fatty acids and chronic stressinduced modulations of glutamatergic neurotransmission in the hippocampus[J]. Nutrition Reviews, 2014, 72(2): 99-112.
[58] RAPOPORT S I, RAO J S, IGARASHI M. Brain metabolism of nutritionally essential polyunsaturated fatty acids depends on both the diet and the liver[J]. Prostaglandins, Leukotrienes, and Essential Fatty Acids, 2007, 77(5/6): 251-261.
[59] RAPOPORT S I. Arachidonic acid and the brain[J]. Journal of Nutrition, 2008, 138(12): 2515-2520.
[60] DEMAR J C, MA K, BELL J M, et al. Half-lives of docosahexaenoic acid in rat brain phospholipids are prolonged by 15 weeks of nutritional deprivation of n-3 polyunsaturated fatty acids[J]. Journal of Neurochemistry, 2004, 91(5): 1125-1137.
[61] BAZAN N G. Omega-3 fatty acids, pro-inflammatory signaling and neuroprotection[J]. Current Opinion in Clinical Nutrition and Metabolic Care, 2007, 10(2): 136-141.
[62] SUN G Y, XU J, JENSEN M D, et al. Phospholipase A2 in the central nervous system: implications for neurodegenerative diseases[J]. Journal of Lipid Research, 2004, 45(2): 205-213.
[63] PIZZA V, AGRESTA A, D’ACUNTO C W, et al. Neuroinflammation and ageing: current theories and an overview of the data[J]. Reviews on Recent Clinical Trials, 2011, 6(3): 189-203.
[64] LITTLE S J, LYNCH M A, MANKU M, et al. Docosahexaenoic acidinduced changes in phospholipids in cortex of young and aged rats: a lipidomic analysis[J]. Prostaglandins, Leukotrienes, and Essential Fatty Acids, 2007, 77(3/4): 155-162.
[65] LATOUR A, GRINTAL B, CHAMPEIL-POTOKAR G, et al. Omega-3 fatty acids deficiency aggravates glutamatergic synapse and astroglial aging in the rat hippocampal CA1[J]. Aging Cell, 2013, 12(1): 76-84.
[66] CHEN C T, GREEN J T, ORR S K, et al. Regulation of brain polyunsaturated fatty acid uptake and turnover[J]. Prostaglandins, Leukotrienes, and Essential Fatty Acids, 2008, 79(3/5): 85-91.
[67] LEE A G. How lipids affect the activities of integral membrane proteins[J]. Biochimica et Biophysica Acta, 2004, 1666(1/2): 62-87.
[68] ZIMMERBERG J, GAWRISCH K. The physical chemistry of biological membranes[J]. Nature Chemical Biology, 2006, 2(11): 564-567.
[69] FAROOQUI A A, HORROCKS L A. Brain phospholipases A2: a perspective on the history[J]. Prostaglandins, Leukotrienes, and Essential Fatty Acids, 2004, 71(3): 161-169.
[70] BAZAN N G. Synaptic signaling by lipids in the life and death of neurons[J]. Molecular Neurobiology, 2005, 31(1/2/3): 219-230.
[71] ULU A, HARRIS T R, MORISSEAU C, et al. Anti-inflammatory effects of omega-3 polyunsaturated fatty acids and soluble epoxide hydrolase inhibitors in angiotensin-II-dependent hypertension[J]. Journal of Cardiovascular Pharmacology, 2013, 62(3): 285-297.
[72] DEPNER C M, TORRES-GONZALEZ M, TRIPATHY S, et al. Menhaden oil decreases high-fat diet-induced markers of hepatic damage, steatosis, inflammation, and fibrosis in obese Ldlr
-/-mice[J]. Journal of Nutrition, 2012, 142(8): 1495-1503.
[73] BANG H O, DYERBERG J, SINCLAIR H M. The composition of the Eskimo food in north western Greenland[J]. American Journal of Clinical Nutrition, 1980, 33(12): 2657-2661.
[74] HOEVENAAR-BLOM M P, NOOYENS A C, KROMHOUT D, et al. Mediterranean style diet and 12-year incidence of cardiovascular diseases: the EPIC-NL cohort study[J]. PLoS ONE, 2012, 7(9): e45458. doi: 10.1371/journal.pone.0045458.
[75] 中国营养学会. 中国居民膳食营养素参考摄入量[J]. 营养学报, 2001, 23(3): 193-196.
Recent Advances in Undersanding the Effects of Polyunsaturated Fatty Acids on Brain Function
LIU Zhiguo, WANG Limei, WANG Hualin, LIU Lieju*
(School of Biology and Pharmaceutical Engineering, Wuhan Polytechnic University, Wuhan 430023, China)
Abstract:Polyunsaturated fatty acids (PUFAs) are a group of important nutritional fatty acids which are involved in cell metabolism and plasma membrane constitution. PUFAs have attracted tremendous attention as an abundant component of brain tissue which plays a crucial role in brain development and improvement of learning and memory abilities. Herein, we summarize the relationship between dietary sources of PUFAs and brain phospholipid composition. Furthermore, the mechanisms of PUFAs on brain development and function are reviewed, which will provide a theoretical basis for the modification of dietary ω-6/ω-3 PUFA ratio in China via adding docosahexaenoic acid (DHA), eicosapentaenoic acid (EPA) or related substitutes from deep-sea fish oil and algae.
Key words:polyunsaturated fatty acid; phospholipid; brain development; brain function
中图分类号:TS201.4
文献标志码:A
文章编号:1002-6630(2015)21-0284-07
doi:10.7506/spkx1002-6630-201521053
收稿日期:2014-12-03
基金项目:国家自然科学基金面上项目(31271855);国家自然科学基金青年科学基金项目(31000772;81402669);
湖北省自然科学基金项目(2014CFB887);湖北省教育厅科学技术研究项目(D20141705)
作者简介:刘志国(1963—),男,教授,博士,研究方向为营养与食品安全。E-mail:zhiguo_l@126.com
*通信作者:刘烈炬(1952—),男,教授,硕士,研究方向为营养生物学。E-mail:liulieju@qq.com