 ±s表示,经SPSS 17.0软件进行单因素方差分析,采用Duncan氏新复极差检验法进行显著性差异分析处理,显著水平P<0.05。
±s表示,经SPSS 17.0软件进行单因素方差分析,采用Duncan氏新复极差检验法进行显著性差异分析处理,显著水平P<0.05。
张 龙,刘俊荣*,田元勇,刘金洋,吴 忠,王 伟
(大连海洋大学食品科学与工程学院,辽宁 大连 116023)
摘 要:从食品原料学角度出发,为系统阐述菲律宾蛤仔的工艺学特性,以含氮物质为出发点,蛋白质为重点,对其各个软体解剖部位的质量组成、一般化学组成、含氮物分布及蛋白质组分等进行了分析讨论。结果表明:菲律宾蛤仔可食部分占原料20%左右,各个软体组织(足、闭壳肌、外套膜、水管、鳃、内脏团)含有水分74.01%~80.07%,其他化学组成为干基,粗蛋白29.19%~43.46%、总糖12.46%~30.75%、粗脂肪1.61%~6.84%和灰分6.69%~11.10%;各组织部位的含氮组分分布比例依次为:非蛋白氮14.7%~26.42%、水溶性蛋白12.55%~19.17%、盐溶性蛋白34.83%~50.4%、碱溶性蛋白17.49%~25.06%及基质蛋白2.19%~10.23%;十二烷基硫酸钠-聚丙烯酰胺凝胶电泳分析显示:菲律宾蛤仔软体部位不同组织、不同组分的蛋白图谱有所差异。水溶性蛋白分子质量较小,在100 kD以下;盐溶性蛋白种类丰富,在200、100、45、35、20 kD附近有明显条带;碱溶性蛋白仅在200、100、45 kD附近有4 条明显条带;基质蛋白分子质量分布范围较宽,主要集中在200 kD以上和45 kD区域。
关键词:菲律宾蛤仔;软体部位;非蛋白氮;蛋白质;分布特性
菲律宾蛤仔(Ruditapes philippinarum)俗称花蛤、蚬子、杂色蛤等,属软体动物门(Mollusca)、双壳纲(Veneroida)、帘蛤科(Veneridae)、蛤仔属(Ruditapes) [1]。广泛分布在我国南北海区,目前是我国四大养殖贝类之一,据2014中国渔业年鉴统计,2013年蛤仔约占我国海水养殖总量的22%,占贝类养殖总量的33% [2]。菲律宾蛤仔是辽宁省最主要的滩涂贝类养殖品种。
作为重要的经济贝类品种,菲律宾蛤仔的食品原料学属性被关注的较少。国内外针对菲律宾蛤仔的研究主要集中在贝类生态学、遗传育种、苗种培育以及养殖技术等方面 [3]。从生物资源利用角度出发,有学者针对菲律宾蛤仔化学组成 [4]及其生物活性成分(如糖蛋白 [5]、溶菌酶 [6]等)的报道,少量加工利用方面(如调味汁 [7]等)的报道。针对经济贝类,国内学者对马氏珠母贝 [8]、波纹巴非蛤 [9]、牡蛎 [10]等,泰国学者对亚洲文蛤 [11]软体部位的含氮物进行了分离分析。
菲律宾蛤仔的可食部分为整个软体组织。本研究以含氮物质为主线,蛋白质为重点,对菲律宾蛤仔的食品原料学属性进行剖析,系统揭示菲律宾蛤仔软体组织的分布特性、含氮物组成和分布、以及蛋白质组成与分布特性。
1.1 原料及处理
活菲律宾蛤仔购自辽宁大连长兴水产品市场(2014 年9—11月),规格为25~30 个/kg。将新鲜原料去壳,除去体腔液,收集可食用部分(软体部位),并用解剖刀分割,包括足、闭壳肌、外套膜、水管、鳃、内脏团,分装标记后,于-18 ℃冻藏备用。
1.2 试剂与仪器
十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate and polyacrylamide gelelectrophoresis,SDSPAGE)试剂 美国Sigma公司;蛋白标准品 美国BioLabs公司;KCl、CuSO 4、NaOH、苯酚 天津市科密欧化学试剂有限公司;Tris 北京索莱宝科技有限公司;三氯乙酸、K 2SO 4、硼酸 国药集团化学试剂有限公司;其他试剂均为分析纯或化学纯。
GL-21M高速冷冻离心机 湖南湘仪实验仪器开发有限公司;721型分光光度计 上海光谱仪器有限公司;MV-Ⅲ垂直平板电泳槽 大连竞迈生物科技有限公司;FJ-200高速分散均质机 上海标本模型厂;BS224S分析天平 北京赛多利斯有限公司;101-1型电热鼓风干燥箱 上海实验仪器厂有限公司;UDK-159全自动凯氏定氮仪 北京盈盛恒泰科技有限责任公司;MS-25制冰机 常州市雪科电器有限公司;PHS-3C精密pH计 上海精密科学有限公司;79-1型磁力搅拌器常州国华电器有限公司。
1.3 方法
1.3.1 含氮物的分离
含氮物的分离参照Hashimoto等 [12]的方法并略有修改。具体如下,所有操作均在4 ℃条件下进行。样品(2.5 g)转移到10 倍体积的0.1 mol/L KCl溶液,20 mmol/L Tris-HCl(pH 7.5)中均质2 min,在磁力搅拌器300 r/min的转速下搅拌30 min。在冷冻离心机12 000×g条件下离心10 min,此过程重复5 次,并收集所有上清液。在所得上清液中加入等量10%三氯乙酸溶液,在12 000×g条件下离心10 min,所得沉淀为水溶性蛋白组分,剩余的上清液为非蛋白氮。
在上述低盐缓冲液洗后的沉淀中加入10 倍体积的0.5 mol/L KCl、20 mmol/L Tris-HCl(pH 7.5)溶液搅拌30 min,然后在12 000×g条件下离心10 min,重复操作4 次,合并所有上清液,并作为盐溶性蛋白组分。在沉淀中加入10 倍体积的0.1 mol/L NaOH溶液,持续搅拌4 h,然后在12 000×g条件下离心10 min,此操作重复4 次。合并的上清液为碱溶性蛋白组分,最终的沉淀为碱不溶性蛋白质,作为基质蛋白组分。
1.3.2 一般组分测定
水分含量的测定:参照GB 5009.3—2010《食品中水分的测定》,采用直接干燥法;粗灰分含量的测定:参照GB 5009.4—2010《食品中灰分的测定》,采用灼烧称重法;粗蛋白质含量的测定:参照GB 5009.5—2010《食品中蛋白质的测定》;粗脂肪含量的测定:参照GB/T 5009.6—2003《食品中脂肪的测定》,采用三氯乙酸沉淀法;总糖含量的测定:参照GB 9695.31—2008《肉制品:总糖含量测定》,采用分光光度法。根据样品的湿质量和干质量对其含量进行计算。
1.3.3 含氮物含量测定
所有含氮组分中氮含量的测定均采用微量凯氏定氮法。各个组织部位的含氮物含量统一以总氮计。
1.3.4 蛋白质组分分子质量的测定
采用SDS-PAGE进行蛋白质组分分子质量的测定,其中浓缩胶质量分数为5%,分离胶质量分数为7.5%,采用考马斯亮蓝R-250染色法检验,醋酸-甲醇溶液进行脱色。
1.4 统计分析
所有实验组重复3 次,使用3 个不同批次的蛤进行实验。数据用
±s表示,经SPSS 17.0软件进行单因素方差分析,采用Duncan氏新复极差检验法进行显著性差异分析处理,显著水平P<0.05。
2.1 菲律宾蛤仔软体部位的组成特性
2.1.1 一般质量组成含量
一般地,经济贝类的可食部位有两大类,一类为部分软体部位可食,如扇贝(仅闭壳肌、外套膜可食);另一类为整体软体部位可食,如牡蛎、蛤仔等。此外,活体贝类体腔液往往构成贝类加工的重要副产物——蒸煮液,富含大量呈味物质 [13]。
表1 菲律宾蛤仔的一般质量组成含量(
 s,n==33)
s,n==33)
Table 1 The general weight composition ofRuditaappeess philpinarum
 s, n == 33)%
s, n == 33)%
部位 贝壳 软体部位 体腔液含量 46.67±1.55 20.49±1.38 32.84±0.17
由表1可知,菲律宾蛤仔软体部位整体可食,占20%左右。辛草等 [14]对3 种经济扇贝的分析,虾夷扇贝、海湾扇贝及栉孔扇贝的软体部位分别占40%、50%、45%,但其主要可食部分为闭壳肌,分别占12.41%、11.24%、14.08%。
菲律宾蛤仔的体腔液占整贝的30%左右,比例较高。从作为民间烹饪的汤料到工业上的煮汁,体腔液以其鲜美的风味吸引着消费者。体腔液的风味功能逐渐引起学者的关注,其化学组成(尤其是呈味物质组成)具有进一步研究与利用空间。对活品贝类来讲,体腔液是反映生命体征的一项指标,杨婷婷 [15]用体腔液体积和pH值评价活品虾夷扇贝的生命体征。
此外,贝壳约占整贝的一半,贝壳含有大量碳酸钙,可以作为良好的钙质饲料。随着贝类养殖业的发展,贝壳将越来越多,为充分利用贝壳资源,现已应用于养殖、医药等方面 [16]。
2.1.2 软体部位组成
菲律宾蛤仔具有整体软体部位可食的特点,可食部分组织结构比较复杂,这不同于以贝柱为主的扇贝,因此,有必要对其软体部位细分后进行研究。软体部位包括足、闭壳肌、外套膜、水管、鳃、内脏团六部分。由表2可知,菲律宾蛤仔各组织所占比例,其中内脏团占软体部位比例最大37%,鳃占软体部位比例最小在9.0%左右,其他组织在11%~15%。
表2 菲律宾蛤仔软体部位的组织结构及质量组成(x ±s,n==33)
Table 2 The structure and weight composition of soft body tissues of Ruditapes philippinarum (x , = 3)

组织 足 闭壳肌 外套膜 水管 鳃 内脏团占整贝比例 2.39±0.20 3.00±0.54 2.63±0.23 3.03±0.24 1.87±0.59 7.58±0.05占软体比例 11.65±0.20 14.62±1.77 12.81±2.19 14.79±0.20 9.13±2.43 37.00±2.43
海洋贝类的风味与含氮物质密不可分,一方面,滋味与非蛋白氮的呈味成分密不可分;另一方面食品的质地与蛋白质密切关联,直接反映到食物的咀嚼感 [17]。因此,有必要对食品原料,特别是动物源食品的可食组织部位的含氮物质进行剖析。
2.2 各组织的一般化学组成分析
菲律宾蛤仔软体部位各组织的一般化学组成见表3。水分含量74.01%~80.07%、粗蛋白含量29.19%~43.46%、总糖含量12.46%~30.75%、粗脂肪含量1.61%~6.84%和灰分含量6.69%~11.10%。吴云霞等 [4]测得菲律宾蛤仔软体部位含量分别为:水分81.49%、粗蛋白61.28%、总糖7.32%、粗脂肪8.61%和灰分17.73%,相比而言,本研究选用的菲律宾蛤仔粗蛋白、粗脂肪和灰分含量略低,总糖含量偏高。随季节、产地等不同,一般化学组成含量会有所不同。
表3 菲律宾蛤仔各组织的一般化学组成含量(x ±s,n==33)
Table 3 Proximate composition of soft tissues ofRuditapes philipnarum (x ± s, n == 33)%
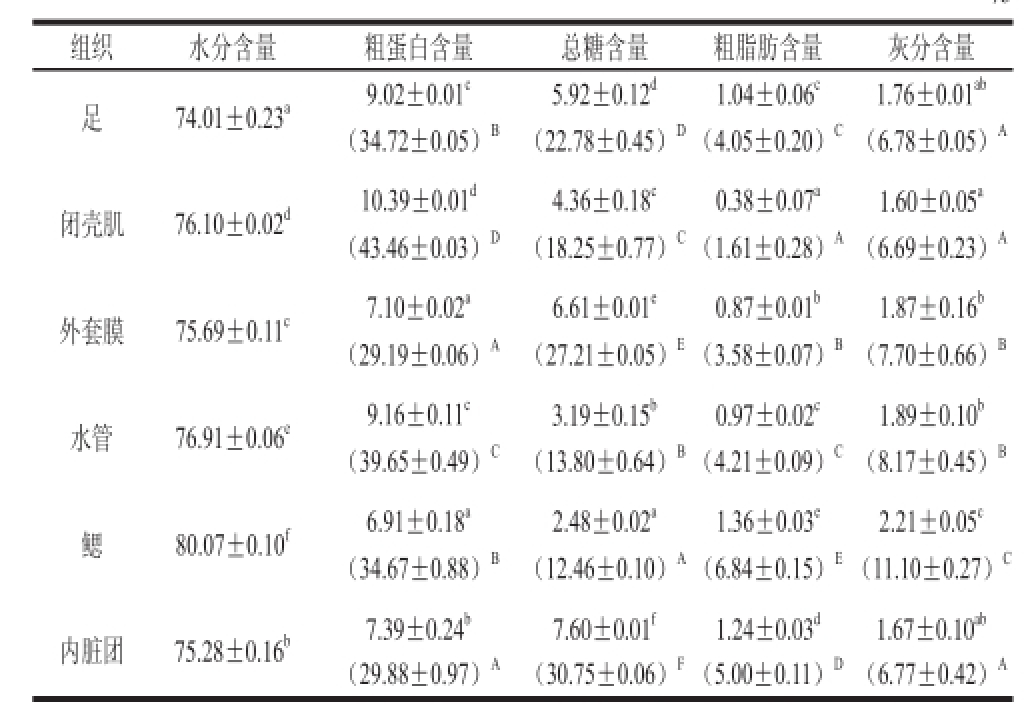
注:表中数据是以湿基计算的含量,括号内数据为以干基计算的含量。小写字母不同表示不同组织(以湿基计算)间差异显著(P<0.05),大写字母不同表示不同组织(以干基计算)间差异显著(P<0.05)。
组织 水分含量 粗蛋白含量 总糖含量 粗脂肪含量 灰分含量足 74.01±0.23 a 9.02±0.01 c 5.92±0.12 d 1.04±0.06 c 1.76±0.01 ab(34.72±0.05) B (22.78±0.45) D(4.05±0.20) C(6.78±0.05) A闭壳肌 76.10±0.02 d 10.39±0.01 d 4.36±0.18 c 0.38±0.07 a 1.60±0.05 a(43.46±0.03) D (18.25±0.77) C(1.61±0.28) A(6.69±0.23) A外套膜 75.69±0.11 c 7.10±0.02 a 6.61±0.01 e 0.87±0.01 b 1.87±0.16 b(2 9.19±0.06) A (27.21±0.05) E(3.58±0.07) B(7.70±0.66) B水管 76.91±0.06 e 9.16±0.11 c 3.19±0.15 b 0.97±0.02 c 1.89±0.10 b(39.65±0.49) C (13.80±0.64) B(4.21±0.09) C(8.17±0.45) B鳃 80.07±0.10 f 6.91±0.18 a 2.48±0.02 a 1.36±0.03 e 2.21±0.05 c(34.67±0.88) B (12.46±0.10) A(6.84±0.15) E(11.10±0.27) C内脏团 75.28±0.16 b 7.39±0.24 b 7.60±0.01 f 1.24±0.03 d 1.67±0.10 ab(29.88±0.97) A (30.75±0.06) F(5.00±0.11) D(6.77±0.42) A
在所有组织中,粗蛋白含量最高的是闭壳肌和水管,分别为43.46%和39.65%,外套膜和内脏团则较低,均在29%左右。与鱼类 [18]相比,贝类中糖原含量高,可见,总糖的含量相对较高,内脏团含量最高约为30%,鳃最低约为12%。一般地,贝类脂肪主要富集在内脏,而肌肉中含量很低,同样,菲律宾蛤的脂肪主要分布在鳃和内脏,分别为6.84%和5.00%,闭壳肌则为1.61%。灰分最高的是鳃11.10%,最低的是闭壳肌6.69%。
不同组织间的一般化学组成含量存在显著性差异,这与各组织在活体中所发挥的功能密切相关 [19]。可见,菲律宾蛤仔各组织中的含氮物为重要组成部分,需进一步针对其各软体组织中含氮物组成及分布进行系统分析。
2.3 各组织含氮物的分离与分析
表 4 菲律宾蛤仔软体部位各组织氮的分布(x ±s,n==33)
Table 4 Nitrogen distribution in soft tissues ofRuditaappeess philipinarmm (x ± s, n == 33)%

注:表中小写字母不同表示不同组织间差异显著(P<0.05)。
组织 非蛋白氮含量 水溶性蛋白含量 盐溶性蛋白含量 碱溶性蛋白含量 基质蛋白含量足 15.64±0.76 a 15.11±2.50 ab 45.33±1.84 b 21.18±0.21 b 4.31±0.24 b闭壳肌 14.70±0.8 a 12.55±1.58 a 50.40±3.74 c 25.06±1.05 c 2.19±0.37 a外套膜 26.42±3.39 c 14.23±1.08 ab 43.17±1.6 b 22.79±1.99 bc 8.42±0.17 c水管 15.87±2.09 a 17.56±4.13 ab 42.77±1.32 b 17.49±0.32 a 7.53±1.22 c鳃 22.79±1.505 b 19.17±2.08 b 34.83±1.29 a 24.19±2.36 bc 8.12±0.27 c内脏团 20.20±0.52 b 15.07±0.27 ab 36.19±1.31 a 18.38±1.08 a 10.23±0.34 d
在食品风味学领域,海产品区别于其他食品原料的特征之一就是,所谓的“鲜”,尤其是海水贝类。由表4可知,非蛋白氮含量为14.7%~26.42%。非蛋白氮或抽提氮中含有游离氨基酸、小肽、核苷酸及其关联物等呈味物质,赋予海产品鲜美的滋味 [20]。各组织含量依次为:外套膜>鳃>内脏团>水管>足>闭壳肌。曾有日本学者根据扇贝闭壳肌浸出物成分(主要是含氮物)分析结果配制出合成浸出物,使之产生贝类特有的风味 [17]。有研究表明,马氏珠母贝肉提取液中游离氨基酸的氮含量占其非蛋白氮的62.8%,且呈味氨基酸含量高,约37%的含氮成分为小肽、核苷酸关联物 [21]。同时,在水产品贮藏加工过程中,发现浸出物成分的化学变化同呈味、鲜度有关。蒋小强等 [22]对冻结后贝肉中非蛋白氮含量的变化进行了研究,杨文鸽等 [23]分析了冰藏期间缢蛏中游离氨基酸、一磷酸腺苷(adenosine monophosphate,AMP)等呈味物质的变化。然而,对菲律宾蛤仔非蛋白氮中的成分(尤其是呈味成分)与含量有待进一步研究。水溶性蛋白(即肌浆蛋白)包括肌红蛋白和大量的酶类,主要是糖酵解酶系,还有磷酸戊糖途径酶系和肌酸激酶、AMP-脱氨酶等 [20],这些蛋白质溶于水或低离子强度的中性盐溶液,是肉中最易提取的蛋白质。表4中水溶性蛋白含量在12.55%~19.17%之间,各组织含量依次为:鳃>水管>足>内脏团>外套膜>闭壳肌。肌浆蛋白与水产品加工及贮藏稳定性有密不可分的关系。Niki等 [24]对扇贝闭壳肌肌浆蛋白凝胶性与其新鲜度的关系进行研究,Kim等 [25]研究表明肌浆蛋白可以改善鱼糜凝胶特性。传统鱼糜凝胶技术将肌浆蛋白通过漂洗脱出,肌浆蛋白的回收成为国内外关注的焦点 [26-32],利用蛋白质等电点发生凝集原理,可以有效分离回收这部分水溶性蛋白。
在食品属性方面,与质地(口感)相关的物性学特征往往由盐溶性蛋白及基质蛋白的质和量所决定。
盐溶性蛋白主要是肌原纤维蛋白组分,许多学者对鱼类肌原纤维蛋白进行研究,以期通过不同的加工参数来改变鱼类肌原纤维蛋白的功能特性 [33],水产动物从采捕到死后肌原纤维蛋白的变化与其品质直接相关。同样,对贝类盐溶性蛋白的研究十分有必要。分析发现(表4),盐溶性蛋白(34.83%~50.4%)为菲律宾蛤仔各个组织的主要含氮物质。各组织含量依次为:闭壳肌>足>外套膜>水管>内脏团>鳃。从现有对贝类的相关研究来看,大部分贝类可食部位均以盐溶性蛋白为主 [9,14],这与本研究结果相符。另外一些种类也有例外,马氏珠母贝 [8]、牡蛎 [10]等以水溶性蛋白为主,亚洲文蛤等 [11]则以碱溶性蛋白为主。
菲律宾蛤仔软体组织中基质蛋白含量2.19%~10.23%(表4),各组织含量依次为:内脏团>外套膜>鳃>水管>足>闭壳肌。基质蛋白包括未溶出的收缩蛋白(如肌联蛋白、肌间线蛋白)、膜蛋白以及大量的结缔组织蛋白 [20]。在肌肉中存在最多的基质蛋白是胶原蛋白,目前,对鱼类胶原蛋白研究很多,对双壳贝类 [1 4,34]中胶原蛋白研究较为缺乏。
菲律宾蛤仔软体组织中碱溶性蛋白含量在17.49%~25.06%(表4),各组织含量依次为:闭壳肌>鳃>外套膜>足>内脏团>水管。Kristinsson等 [35]称其主要成分为交联的肌原纤维蛋白,还可能存在变性的肌动球蛋白和少量的基质蛋白 [12]。Karnjanapratum等 [11]认为,根据各组织的功能,交联蛋白质可以加强相应组织的强度。
不同组织中的含氮物分布存在显著差异,这种差异与各组织的结构、功能密不可分。在足和闭壳肌中,高含量的盐溶性蛋白组分和碱溶性蛋白组分可能有助于形成其坚实的质地,在外套膜、水管、鳃和内脏团中,高含量的基质蛋白组分可能与其较高的韧性有关。
2.4 各组织的蛋白质组分分析

图1 菲律宾蛤仔软体部位各组织的SDS-PAGEE图谱
Fig.1 SDS-PAGE patterns of proteins in soft tissues of Ruditapes philippinarum
如图1所示,菲律宾蛤仔不同组织、不同溶解组分的SDS-PAGE图谱有所差异。首先,比较各组织的原料肉条带,可见,足、闭壳肌、外套膜、水管中的蛋白组成相似,蛋白种类丰富。收缩蛋白组分(粗、细丝蛋白)清晰可见,此外,在200 kD以上还出现一条明显条带(箭头所示),可能含肌球蛋白重链(myosin heavy chain,MHC)二/多聚体 [36]、颤搐蛋白(twitchin,TW)、胶原蛋白等。然而,鳃和内脏团中的蛋白组成相似,蛋白种类相对较少,分子质量主要分布在60 kD以下。
对于各组织中的水溶性蛋白来说,足、闭壳肌、外套膜、水管、鳃、内脏团的分子质量分别分布为<60 kD、<100 kD、<60 kD、<60 kD、<35 kD、<45 kD。足、闭壳肌、外套膜、水管的蛋白条带比鳃和内脏的分布范围要广。其中较明显条带出现在:足(约45、35、28 kD)、闭壳肌(约45、24 kD)、外套膜(约30、35 kD)、水管(约30、24 kD)。除闭壳肌外,其他部位均在20 kD以下出现鲜明涂抹带。
盐溶性蛋白所含蛋白种类是最丰富的,主要成分为肌原纤维蛋白(粗、细丝)。软体动物的粗丝蛋白由作为核心的副肌球蛋白(paramyoosin,PM),以及肌球蛋白(myosin,MY)、Myorod(MR) [37-39]、TW [39-46]组成;细丝蛋白包括肌动蛋白(actin,AC)、原肌球蛋白、肌钙蛋白或钙调节蛋白 [47-48]。除鳃以外的各组织中均出现明显条带:约200 kD的MHC、约100 kD的副肌球蛋白、约45 kD的肌动蛋白或钙调节蛋白、约35 kD的原肌球蛋白、小于20 kD的肌球蛋白轻链。此外,还出现了位于MHC之上的未知条带(箭头所示),很可能是TW,现有研究发现,在贻贝(闭壳肌和外套膜)、虾夷扇贝(闭壳肌外侧的平滑肌)、牡蛎(闭壳肌)中存在TW,分子质量约600 kD [39-46]。除鳃和内脏团外,还可能存在MR的条带,其由MHC基因产生,贻贝(Mytilus)中含2 条分子质量在115~130 kD的多肽链 [37],有报道称其只存在于软体动物平滑肌中 [42]。现对贝类来说,在贻贝、牡蛎和扇贝的平滑肌中存在MR。
碱溶性蛋白中蛋白种类简单,在足、闭壳肌、外套膜、水管中均出现明显的4 条条带,分子质量约在200、100、45 kD以及高于200 kD。而鳃和内脏团中的条带微乎其微。“TW-AC联动假说”中指出软体动物平滑肌放松极缓慢,消耗能量极低(所谓的“闭壳现象”)与TW-AC联动机制有密切关系。明确指出交联的肌原纤维蛋白存在形式有:TW-MY、TW-AC、PM-TW-MY等 [40-42],因此推测碱溶性蛋白成分主要是由MY、PM、AC、TW形成的交联蛋白,这4种蛋白组分正好对应于电泳图谱中的4 条条带。
除鳃外,其他组织的基质蛋白分子质量主要集中在200 kD以上和45 kD,这表明交联结缔组织蛋白的存在。闭壳肌中还在100、35 kD上下分别出现2 条明显条带,鳃中无明显条带。在肌肉中存在最多的基质蛋白就是胶原蛋白,肌肉组织的韧性主要与胶原蛋白有关。200 kD以上的条带可能为大量的胶原蛋白β二聚体、γ三聚体,其中,β链由2 条分子质量为100 kD的α链通过氢键连接而成,γ链由3 条分子质量为100 kD的α链通过氢键连接而成 [20]。关于双壳贝类胶原蛋白研究较少,辛草等 [14]发现虾夷扇贝、海湾扇贝、栉孔扇贝的外套膜胶原蛋白条带主要集中分布在212~185 kD,为β肽链,而α肽链不明显。
从食品原料学角度研究经济贝类是非常必要的。丰富的非蛋白氮和蛋白氮赋予贝类特有的滋味和口感,因此,含氮物可以从整体上来诠释其重要的食品属性(风味、质地)。菲律宾蛤仔的可食部位包含整个软体部位,主要由足、闭壳肌、外套膜、水管、鳃以及内脏团6 个组织构成。从含氮物的分析表明,这些不同软体组
织间存在差异,这与各部位的结构、功能密不可分。值得注意的是菲律宾蛤仔中含有某些贝类所特有的蛋白质(TW和MR),在下一步的研究中将进行更深入地验证与探讨。本研究从食品原料学特性出发,系统探索了菲律宾蛤仔可食部位的含氮物组分及分布特性,该研究对于指导采捕后流通过程中的品质管理、以及贝类产品研发具有实践参考价值。
参考文献:
[1] 张国范, 闫喜武. 蛤仔养殖学[M]. 北京: 科学出版社, 2009: 1.
[2] 农业部渔业局. 中国渔业统计年鉴[M]. 北京: 中国农业出版社,2014: 32-33.
[3] 刘青, 张越, 付鑫, 等. 菲律宾蛤仔的研究进展[J]. 河北渔业, 2011(1):56-59.
[4] 吴云霞, 梁健, 闫喜武, 等. 菲律宾蛤仔营养成分分析与评价[J]. 营养学报, 2012, 34(4): 409-410.
[5] 吴红棉, 叶志国, 范秀萍, 等. 菲律宾蛤仔糖蛋白的分离纯化与理化性质的研究[J]. 中国食品学报, 2009, 9(5): 80-85.
[6] KIM M, PARK M, JEONG Y. Purification and characterization of lysozyme from filipino venus, Ruditapes philippinarum[J]. Food Science and Biotechnology, 2012, 21(5): 1463-1468.
[7] 林金璇. 菲律宾蛤仔调味汁工艺及其流变性质研究[D]. 福州: 福建农林大学, 2012.
[8] 郑惠娜, 章超桦, 刘书成, 等. 马氏珠母贝蛋白的分离及分子量分布研究[J]. 水产科学, 2008, 27(9): 447-451.
[9] 何小庆, 曹文红, 赵子科, 等. 波纹巴非蛤蛋白分离及其性质研究[J].食品与发酵工业, 2013, 39(5): 229-233.
[10] 张晶晶, 郑惠娜, 章超桦, 等. 牡蛎蛋白分离及其基本组成分析[J].食品与发酵工业, 2013, 39(9): 195-199.
[11] KARNJANAPRATUM S, BENJAKUL S, KISHIMURA H, et al. Chemical compositions and nutritional value of Asian hard clam (Meretrix lusoria) from the coast of Andaman Sea[J]. Food Chemistry,2013, 141(4): 4138-4145.
[12] HASHIMOTO K, WATABE S, KONO M, et al. Muscle protein composition of sardine and mackerel[J]. Bulletin of the Japanese Society of Scientifi c Fisheries, 1979, 45(11): 1435-1441.
[13] 何炘. 贻贝蒸煮液调味品香气成分及其酿造工艺的研究[D]. 杭州:浙江工商大学, 2007.
[14] 辛草, 刘俊荣, 蔡秋凤, 等. 三种经济扇贝的基本工艺性质及蛋白质分析[J]. 水产科学, 2012, 31(9): 535-539.
[15] 杨婷婷. 采捕后活品虾夷扇贝(Patinopecten yessoensis)的风味变化[D].大连: 大连海洋大学, 2014: 52-55.
[16] 李金志. 贝壳的综合利用[J]. 淮海工学院学报, 2009(10): 22-23.
[17] 鸿巢章二, 桥本周久. 水产利用化学[M]. 北京: 中国农业出版社,1994: 126-127.
[18] SCHULZ M, LIESE A D, MAYER-DAVIS E J, et al. Nutritional correlates of dietary glycaemic index: new aspects from a population perspective[J]. British Journal of Nutrition, 2005, 94(3): 397-406.
[19] GOSLING E. Bivalve molluscs: biology, ecology and culture[M]. US:John Wiley & Sons, 2008.
[20] FOEGEDING E A, LANIER T C, HULTIN H O. Characteristics of edible muscle tissues[J]. Food Chemistry, 1996, 3(15): 879-942.
[21] 章超桦, 吴红棉, 洪鹏志, 等. 马氏珠母贝肉的营养成分及其游离氨基酸组成[J]. 水产学报, 2000, 24(2): 180-184.
[22] 蒋小强, 李敏, 关志强. 文蛤和波纹巴非蛤冷冻变性的比较研究[J].南方水产科学, 2006, 2(4): 7-12.
23] 杨文鸽, 徐大伦, 孙翠玲, 等. 缢蛏冰藏保活期间呈味物质的变化[J].中国食品学报, 2009, 9(3): 181-186.
24] NIKI T, KATO Y, NOZAWA H, et al. Gelation of low salt soluble proteins from scallop adductor muscle in relation to its freshness[J]. Fisheries Science, 2002, 68(3): 688-693.
25] KIM Y S, YONGSAWATDIGUL J, PARK J W, et al. Characteristics of sarcoplasmic proteins and their interaction with myofibrillar proteins[J]. Journal of Food Biochemistry, 2005, 29(5): 517-532.
26] 刘诗长. 罗非鱼分离蛋白的制备及其性质研究[D]. 湛江: 广东海洋大学, 2011.
27] 梁姗姗, 刘俊荣, 马永生, 等. 虾夷扇贝(Patinopecten yessoensis)外套膜蛋白的分离提取及功能特性[J]. 食品科学, 2014, 35(7): 12-16. doi: 10.7506/spkx1002-6630-201407003.
28] 闫瑞霞, 刘俊荣, 马永生, 等. 箭齿鲽肌肉分离蛋白功能特性的研究[J].大连海洋大学学报, 2013, 28(5): 492-497.
29] 吴忠, 刘俊荣, 马永生, 等. 鱼蛋白的浓缩分离及其凝胶性的研究进展[J]. 大连海洋大学学报, 2014, 29(5): 536-542.
30] 刘俊荣, 贺凌, 汪涛, 等. 不同pH值条件下罗非鱼肌肉蛋白分离效果及其功能特性的研究[J]. 水产科学, 2010, 29(6): 317-320.
31] 傅润泽, 刘俊荣, 王帅, 等. 采用酸碱提取法对鲢肌肉蛋白分离特性的影响[J]. 大连海洋大学学报, 2012, 26(6): 539-543.
32] 李芳, 刘俊荣, 梁姗姗, 等. 南极磷虾蛋白质的分离特性及其组分分析[J]. 大连海洋大学学报, 2013, 28(2): 191-194.
33] SIKORSKI Z E. The myofibrillar proteins in seafoods[M]//Seafood Proteins. Springer US, 1995: 40-57.
34] MIZUTA S, MIYAGI T, NISHIMIYA T, et al. Partial characterization of collagen in mantle and adductor of pearl oyster (Pinctada fucata)[J]. Food Chemistry, 2002, 79(3): 319-325.
35] KRISTINSSON H G, THEODORE A E, DEMIR N, et al. A comparative study between acid- and alkali- aided processing and surimi processing for the recovery of proteins from channel catfish muscle[J]. Journal of Food Science, 2005, 70(4): 298-306.
36] YOSHIDA W, KUNIMI O, FUJIURA M, et al. Thermal gelation of salted paste from scallop striated adductor muscle[J]. Fisheries Science, 2003, 69(5): 1017-1025.
37] SHELUD'KO N S, PERMJAKOVA T V, TUTUROVA K, et al. Myorod, a thick filament protein in molluscan smooth muscles: isolation, polymerization and interaction with myosin[J]. Journal of Muscle Research and Cell Motility, 2001, 22(1): 91-100.
38] SHELUD'KO N S, TUTUROVA K F, PERMYAKOVA T V, et al. A novel thick fi lament protein in smooth muscles of bivalve molluscs[J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 1999, 122(3): 277-285.
[39] BÉJAR P, VILLAMARÍN J A. Catalytic subunit of cAMP-dependent protein kinase from a catch muscle of the bivalve mollusk (Mytilus galloprovincialis): purifi cation, characterization, and phosphorylation of muscle proteins[J]. Archives of Biochemistry and Biophysics, 2006,450(2): 133-140.
[40] SHELUD'KO N S, MATUSOVSKY O S, PERMYAKOVA T V,et al. “Twitchin-actin linkage hypothesis” for the catch mechanism in molluscan muscles: evidence that twitchin interacts with myosin,myorod, and paramyosin core and affects properties of actomyosin[J]. Archives of Biochemistry and Biophysics, 2007, 466(1): 125-135.
[41] SHELUD'KO N S, MATUSOVSKAYA G G, PERMYAKOVA T V,et al. Twitchin, a thick-fi lament protein from molluscan catch muscle,interacts with F-actin in a phosphorylat ion-dependent way[J]. Archives of Biochemistry and Biophysics, 2004, 432(2): 269-277.
[42] SOBIESZEK A, MATUSOVSKY O S, PERMYAKOVA T V, et al. Phosphorylation of myorod (catchin) by kinases tightly associated to molluscan and vertebrate smooth muscle myosins[J]. Archives of Biochemistry and Biophysics, 2006, 454(2): 197-205.
[43] TSUTSUI Y, YOSHIO M, OIWA K, et al. Striated muscle twitchin of bivalves has “catchability”, the ability to bind thick fi laments tightly to thin fi laments, representing the catch state[J]. Journal of Bolecular Biology, 2007, 365(2): 325-332.
[44] AVROVA S V, SHELUD'KO N S, BOROVIKOV Y S. A new property of twitchin to restrict the “rolling” of mussel tropomyosin and decrease its affi nity for actin during the actomyosin ATPase cycle[J]. Biochemical and Biophysical Besearch Bommunications, 2010,394(1): 126-129.
[45] MATUSOVSKY O S, SHELUD'KO N S, PERMYAKOVA T V, et al. Catch muscle of bivalve molluscs contains myosin-and twitchin-associated protein kinase phosphorylating myorod[J]. Biochimica et Biophysica Acta (BBA)-Proteins and Proteomics, 2010, 1804(4): 884-890.
[46] BOROVIKOV Y S, SHELUD'KO N S, AVROVA S V. Molluscan twitchin can control actin-myosin interaction during ATPase cycle[J]. Archives of Biochemistry and Biophysics, 2010, 495(2): 122-128.
[47] FUNABARA D, NAKAYA M, WATABE S. Isolation and characterization of a novel 45 kDa calponin-like protein from anterior byssus retractor muscle of the mussel Mytilus galloprovincialis[J]. Fisheries Science, 2001, 67(3): 511-517.
[48] DOBRZHANSKAYA A V, MATUSOVSKAYA G G, MATUSOVSKY O S, et al. Thin filaments of bivalve smooth muscle may contain a calponin-like protein[J]. Biophysics, 2010, 55(5): 703-706.
Composition and Distribution of Nitrogenous Compounds in Soft Body of Ruditapes philippinarum
ZHANG Long, LIU Junrong*, TIAN Yuanyong, LIU Jinyang, WU Zhong, WANG Wei
(College of Food Science and Engineering, Dalian Ocean University, Dalian 116023, China)
Abstract:This study investigated quality parameters of soft body tissues of the clam Ruditapes philippinarum such as anatomic composition, proximate composition, and the distribution of nitrogenous compounds with emphasis on proteins. The results were indicated that 1) total edible portions accounted for about 20% of Ruditapes philippinarum. Proximate composition of the edible portions including six tissues, foot, adductor, mantle, siphon, gill and viscera, consisted of moisture (74.01%-80.07%), protein (29.19%-43.46%, dry basis), carbohydrate (12.46%-30.75%, dry basis), fat (1.61%-6.84%, dry basis) and ash (6.69%-11.10%, dry basis); 2) the nitrogenous components were distributed in non-protein nitrogen (14.7%-26.42%), water-soluble protein (12.55%-19.17%), salt-soluble protein (34.83%-50.4%), and alkali-soluble protein (17.49%-25.06%); and 3) sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) analysis showed that the protein patterns varied among the soft tissues and components, with the water-soluble proteins, salt-soluble proteins and alkalisoluble proteins distributed below 100 kD, near 200, 100, 45, 35 and 20 kD, and near 200, 100 and 45 kD, respectively, and the stroma proteins distributed in a wide range, mainly near 45 kD and at 200 kD.
Key words:Ruditapes philippinarum; soft tissues; non-protein nitrogen; protein; composition profi les
中图分类号:TS254
文献标志码:A
文章编号:1002-6630(2015)24-0095-06
doi:10.7506/spkx1002-6630-201524016
收稿日期:2015-03-09
基金项目:国家自然科学基金面上项目(31271980)
作者简介:张龙(1989—),男,硕士研究生,研究方向为水产品加工及贮藏工程。E-mail:shipinzl@126.com
*通信作者:刘俊荣(1963—),女,教授,博士,研究方向为水产品加工。E-mail:ljunrong@dlou.edu.cn