
陈 敏 1,高云慨 1,宋海超 1,何书婷 1,张荣意 1,钟利文 2,史学群 3,*
(1.海南大学环境与植物保护学院,海南 海口 570228;2.海南大学园艺园林学院,海南 海口 5702 28;3.海南大学食品学院,海南 海口 570228)
摘 要:研究氯化 钙(CaCl 2)结合季也蒙毕赤酵母(Meyerozyma guilliermondii)处理对由炭疽病菌(Colletotrichum gloeosporioides)引起的芒果采后炭疽病的抑制效果,同时分 析CaCl 2对病原菌和酵母拮抗菌生长以及果实抗病性的影响。结果显示:40 g/L CaCl 2可有效提升拮抗菌M. g uilliermondii对芒果采后炭疽病的生防效果,其中40 g/L CaCl 2与1×10 8CFU/mL M. guilliermondii悬浮液的复合处理比单一处理更能有效地抑制芒果果实采后炭疽病的发生。此外,40 g/L CaCl 2对C. gloeosporioides菌丝 的生长具有抑制作用,并能够促进拮抗菌M. guilliermondii在芒果果实伤口处的增殖。与此同时,该复合处理显著诱导了芒果果实抗病相关酶活性的升高。结果表明,40 g/L CaCl 2可以直接通过抑制病原菌的生长和促进拮抗酵母的增殖、以及间接诱导果实抗病 性来提升拮抗菌M. guilliermondii对芒果果实采后炭疽病防治效果。
关键词:芒果;炭疽菌;氯化钙;季也蒙毕赤酵母;生防;酶活性
芒果(Mangifera indica Linn.)作为世界五大热带水果之一,以其独特的口感和丰富的营养价值,被誉为“热带水果之王” [1]。芒果炭疽病是世界性的芒果重要病害,其主要病原菌为胞炭疽菌(Colletotrchum gloeosporioides (Penz.) Penz.and Sacc.) [2]。使用化学杀菌剂仍然是该 病采后防治的常用措施 [3-4]。然而,长期使用化学杀菌剂不但会导致病原菌抗药性的产生从而降低化学药剂的防治效果,而且经常使用高浓度化学药剂会导致果实上农药残留量的增加而威胁人类的健康,并造成环境的污染 [5-7]。因此,寻求安全、经济、有效防治果实采后病害的新技术已成为当务之急。近年来,生物拮抗菌作为一种具有潜力的新兴技术,在采后病害防治方面显示出的有 效性和使用方便性广受关注 [8]。
目前,一些酵母拮抗剂已经被广泛应用于水果采后病害控制的研究并具有明显的抑制效果。笔者之前从中国海南昌江地区芒果园筛选到了1 株酵母,经中国普通微生物菌种保藏管理中心(China General Microbiological Culture Collection Center,CGMCC)鉴定为Meyerozyma guilliermondii。实验表明,拮抗菌M. guilliermondii对芒果采后炭疽病具有较好的抑制效果。然而,该拮抗菌单独使用时需要在高浓度条件下才能达到理想的防治效果,其防效与化学杀菌剂相比也存在一定的差距,这些因素极大地限制了该拮抗酵母的大规模商业化应用 [5]。 因此,需要通过其他方法来提高该拮抗菌的生物防治能力。
前人的研究表明,很多外源化学物质,如壳聚糖 [9]、钼酸铵 [10]、碳酸氢钠 [11]、微波 [12]、水杨酸 [13]、热处理 [14]和硅 [10]等与拮抗酵母 结合使用,均能够显著提高拮抗酵母的生物防治活性。
氯化钙(CaCl 2)作为防腐剂和固化剂在果蔬加工行业中已被广泛应用 [15]。之前已有研究表明,CaCl 2可以增强Cryptococcus laurentii对苹果绿霉病和梨灰霉病的防治效果 [16-17]。但 对于CaCl 2是否也能够增强拮抗菌M. guilliermondii对芒果采后炭疽病的生防效果尚未见报道。因此,本实验在先期研究的基础上,进一步探讨了使用CaCl 2增强M. guilliermondii生防效力的可行性,同时对可能涉及到的机理进行探索,旨在为芒果采后炭疽病的生物防治提供理论依据。
1.1 材料
实验所用的芒果品种为“红金龙”,购自海南昌江芒果园,果实成熟度为7~8 成、大小均一、无病虫害,使用前用20 g/L次氯酸钠溶液消毒2 min后用自来水冲洗,自然晾干后备用。
1.2 试剂及培养基
CaCl 2(分析纯) 广州化学试剂公司。
营养酵母-葡萄糖液体培养基:营养肉汤8 g/L、酵母粉5 g/L、葡萄糖10 g/L、氯化钠20 g/L,121 ℃灭菌20 min,待用;马铃薯-葡萄糖(pota to dextrose agar,PDA)培养基:去皮马铃薯200 g/L、葡萄糖20 g/L、琼脂20 g/L,121 ℃灭菌20 min,待用。
1.3 方法
1.3.1 病原菌和拮抗菌
1.3.1.1 病原菌
实验所用的芒果采后炭疽病菌为实验室分离保存的菌株,前期已经单孢分离和分子鉴定为C. gloeosporioides Penz. and Sacc,致病性测定表明为强致病力菌株。将其接种于PDA培养基上,28 ℃条件下培养7 d,孢子悬浮液的制备用含有0.05%吐温-80的无菌水配制成实验所需要浓度,待用。
1.3.1.2 拮抗菌
实验用拮抗酵母分离自海南昌江地区芒果园芒果果实伤口,经CGMCC鉴定为M. guilliermondii。将该酵母菌株接种于含有50 mL营养酵母-葡萄糖液体培养基中,在2 8 ℃条件下振 荡培养48 h,培养液在8 000 r/min条件下离心10 min后,弃上清液,加入无菌水,最后用血球计数板配制成实验所需要的浓度。
1.3.2 CaCl 2对C. gloeosporioides菌丝生长的影响
用无菌打孔器(直径为5 mm)在培养7 d左右的PDA培养基上取炭疽菌菌落边缘的菌块放置于含有不同质量浓度CaC l 2(0(对照)、5、10、20、40、50 g/L的PDA培养基平板(直径为9 cm)中心。培养皿置于28 ℃条件下培养4 d,调查菌落直径的大小。每个处理重复3 次,实验重复2 次。
1.3.3 CaCl 2对拮抗菌M. guilliermondii在果实伤口生长动态的影响
参照Janisiewicz等 [18]的方法进行稍加改进,测定不同质量浓度CaCl 2对拮抗菌M. guilliermondii在果实伤口处的生长动态的影响。分别用质量浓度为0(对照)、5、10、20、40、50 g/L的CaCl 2溶液配制成浓度均为1×10 8CFU/mL的拮抗菌M. guilliermondii悬浮液,各取20 μL接种到芒果果实伤口内(直径为5 mm,深3 mm,以下同),将果实28 ℃条件下保湿贮藏,分别测定0(接种后1 h)、24、48、72、96 h拮抗菌在果实伤口处的生长情况。测定方法为用灭过菌的打孔器从伤口处取直径和高度都为10 mm的果肉组织,放在已加入1.0 mL 0.05 mol/L(pH 6.5)的磷酸缓冲液的研钵中,研磨后采用稀释平 板 法测定拮抗菌的数目。每处理5 个果实,整个实验重复2 次。
1.3.4 CaCl 2与拮抗菌M. guilliermondii配合使用对芒果采后炭疽病的抑制效果影响
每个处理选取7 个芒果果实置于塑料筐中,用打孔器在果实赤道部位制造伤口。芒果果实分别做以下处理:A:40 g/L CaCl 2浸泡10 min;B:伤口处滴加30 μL 1×10 8CFU/mL拮抗菌M. guilliermondii悬浮液;C:40 g/L CaCl 2浸泡10 min后伤口处滴加30 μL 1×10 8CFU/mL拮抗菌M. guilliermondii悬浮液;D:对照为无菌水浸泡10 min。4 h后,各处理果实伤口接入15 μL浓度为5×10 4CFU/mL炭疽孢子悬浮液。果实晾干后用保鲜膜封口以保持湿度(95%左右)28 ℃贮藏,分别于第4、6天测量各果实的病斑直径及发病率。每个处理3 个重复,实验重复2 次。
1.3.5 CaCl 2不同处理时间对芒果采后炭疽病抑制效果的影响
按1.3.4节中的方法对芒果果实制造伤口。各果实分为2 组进行不同处理:A:40 g/L CaCl 2浸泡10 min,2 h后再接种15 μL浓度为5×10 4CFU/mL C. gloeosporioides孢子悬浮液;B:40 g/L CaCl 2浸泡10 min,24 h后再接种15 μL浓度为5×10 4CFU/mL C. gloeosporioides孢子悬浮液。各处理果实28 ℃条件下放置于95%相对湿度的塑料箱中,4 d后分别测定各果实的病斑直径和发病率。每个处理5 个芒果果实,3 次重复,整个实验重复2 次。
1.3.6 CaCl 2和拮抗菌M. guilliermondii处理对芒果果实苯丙氨酸解氨酶(phenylalanineammonialyase,PAL)、过氧化物酶(peroxidase,POD)和多酚氧化酶(polyphenloxidase,PPO)活性的影响
1.3.6.1 酶活性的诱导
每个芒果果实用打孔器在果实赤道部位制造伤口(5 mm×3 mm)。实验分为4 组不同处理:A:刺伤后滴加30 μL无菌水作为对照;B:刺伤后滴加30 μL 40 g/L CaCl 2溶液;C:刺伤后滴加30 μL 1×10 8CFU/mL M. guillierm ondii悬浮液;D:刺伤后滴加40 g/L CaCl 230 μL,10 min后继续加入30 μL、1 ×10 8CFU/mL M. guilliermondii悬浮液。将处理的果实分别贮藏于28 ℃,每24 h取样测定酶活性连续测定5 d。每个处理3 个重复,每个处理5 个果实,实验重复2 次。
1.3.6.2 PAL酶液的制备与活性测定
5 g果肉中加入0.3 g聚乙烯吡咯烷酮,再加入0.05 mol/L的硼酸缓冲溶液25 mL(pH 8.8,含有0.005 mol/L巯基乙醇溶液),冰浴匀浆,12 000 r/min,4 ℃条件下离心45 min,上清液用于酶活性测定。
PAL 活性测定参照Assis等 [19]的方法加以改进:5 mL反应体系中含有1 mL粗酶液、2 mL硼酸缓冲液和1 mL 0.02 mol/L L-苯丙氨酸,37 ℃水浴60 min后,立即加入0.5 mL 6 mol/L HCl溶液终止反应,随后测定A 290 nm值,以反 应液每小时在 290 nm波长处吸光度增加0.01为1 个酶活力单位。酶活性结果以U/mg表示。
1.3.6.3 PPO及POD酶液的制备与活性测定
取5 g果肉组织,加入0.3 g聚乙烯吡咯烷酮,再加入0.1 mol/L的磷酸缓冲液30 mL(pH 6.4),冰浴匀浆,12 000 r/min,4 ℃条件下离心30 min,上清液用于活性测定。
POD活性测定参照Chance等 [20]的方法加以改进:在2 mL 8 mmol/L愈创木酚溶液中(用0.1 mo/L pH 6.4的磷酸钠缓冲液配制成)中加入0.5 mL酶液,于37 ℃水浴5 min后,加入1 mL H 2O 2(24 mmol/L)混匀后测定A 470 nm值,以反应液每分钟在 470 nm波长处吸光度增加0.01 为1 个酶活力单位。酶活性结果以U/mg表示。
PPO活性测定参照文献[21]的方法加以改进:在2.5 mL 50 mmol/L邻苯二酚溶液中(用0.1mol/L pH 6.4的磷酸钠缓冲液配制成)中加入0.5 mL酶液,混合物混匀后置于30 ℃条件下充分5 min,随后测定A 398 nm值,以反应液每分钟在398 nm 波长处吸光度增加0.01为1 个酶活力单位。酶活性结果以U/mg表示。
1.3.6.4 酶液蛋白含量 的测定
参照Brodford [22]的方法进行,以牛血清白蛋白作为标准蛋白。
1.4 数据分析
采用SAS软件进行数据统计,采用ANOVA对数据进行邓肯多重差异分析(P<0.05)。
2.1 CaCl 2对芒果炭疽病菌菌丝扩展的影响
通过研究不同质量浓度(0、5、10、20、40、50 g/L)CaCl 2对C. gloeosporioides在PDA固体培养基上生长的影响,结果见图1,在28 ℃条件下培养4 d后,不同质量浓度的CaCl 2对C. gloeosporioides在PDA培养基上的生长具有一定的影响。其中,质量浓度为5、10、20 g/L的CaCl 2对C. gloeosporioides菌丝的生长具有一定的促进作用,而质量浓度为40 g/L和50 g/L的CaCl 2对C. gloeosporioides菌丝的生长与对照相比则具有一定的抑制作用。

图1 不同质量浓度CaCl
2对芒果炭疽病菌菌丝扩展的抑制效果
Fig.1 Inhibitory effect of different concentrations of calcium chloride on mycelial growth of C. gloeosporioides
A.接种第4天抑制效果图;B.培养基效果图。1~6分别为对照、 5、10、20、40、50 g/L。柱形上不同小写字母表示差异显著(P<0.05),下同。
2.2 CaCl 2对拮抗菌M. guilliermondii在果实伤口生长动态的影响

图2 CaaCCll
2对拮抗菌M. guilliermonnddiiii在芒果果实伤口处生长动态的影响
Fig.2 Effect of different concentrations of calcium chloride on population growth of M. guilliermondii in fruit wounds
分别将质量浓度为0、5、10、20、40、50 g/L的CaCl 2配制的拮抗菌M. guilliermondii悬浮液接种于芒果果实伤口,观察其在芒果伤口的生长动态,结果见图2。从图2可以看出,不同质量浓度CaCl 2配制的拮抗菌悬浮液在芒果果实伤口处都能迅速繁殖。在0~48 h期间,芒果果实伤口处各拮抗菌数量大约是接种时的57~127 倍,随后基本保持稳定。其中,质量浓度为20 g/L和40 g/L的CaCl 2对拮抗菌M. guilliermondii在芒果果实伤口处的生长具有促进作用,而质量浓度为50 g/L的CaCl 2对拮抗菌M. guilliermondii在芒果果实伤口处的生长与对照相比则有轻微的抑制作用。
2.3 CaCl 2和拮抗菌M. guilliermondii结合使用对芒果采后炭疽病抑制效果的影响
将质量浓度为40 g/L CaCl 2与拮抗菌M. guilliermondii结合处理芒果果实,观察40 g/L CaCl 2对拮抗菌M. guilliermondii抑制芒果采后炭疽病的影响,其结果见图3。与对照相比,40 g/L CaCl 2单独使用时对芒果采后炭疽病没有显著的抑制作用,而拮抗菌M. guilliermondii单独使用或结合40 g/L CaCl 2使用能显著降低芒果采后炭疽病病斑直径和发病率。当40 g/L CaCl 2和1×10 8CFU/mL拮抗菌M. guilliermondii悬浮液结合使用时,芒果采后炭疽病病斑直径和发病率比拮抗菌M. guilliermondii单独时有所降低,其中,芒果果实在接种第4天能完全抑制芒果采后炭疽病的发生,尽管接种第6天发病率有所上升但病斑直径仍然显著低于对照。
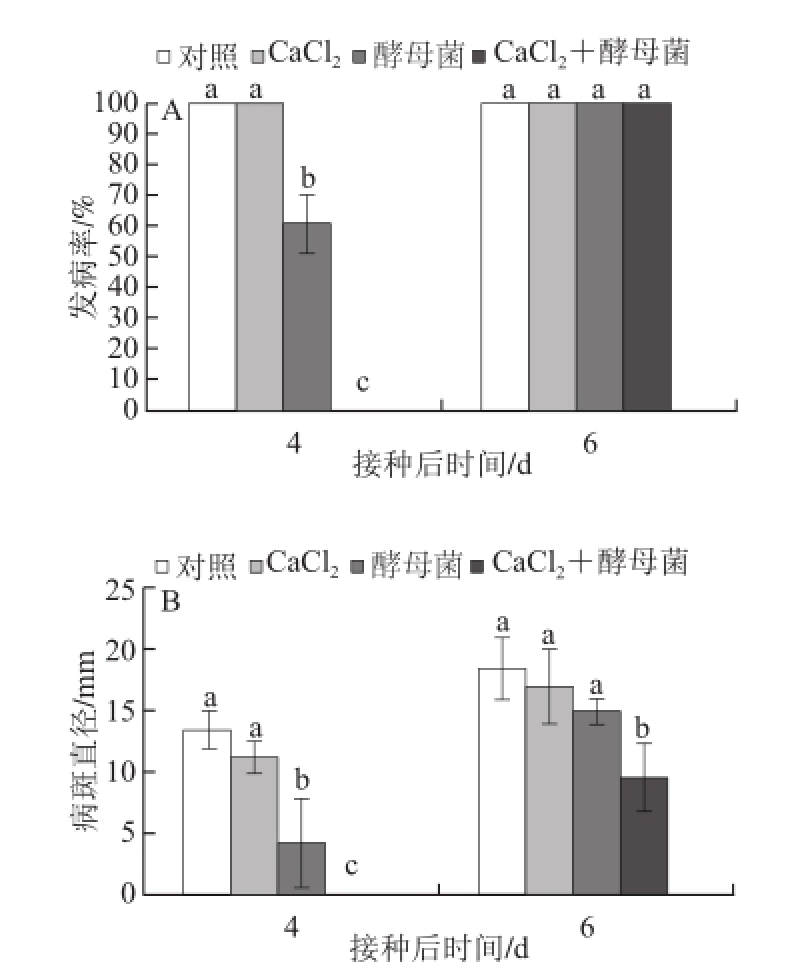
图3 CaCl
2与M. guilliermondii 处理对芒果采后炭疽病发病率(A)和病斑直径(B)的影响
Fig.3 Effects of disease incidence (A) and lesion diameter (B) of C. gloeosporioides in mango fruit postharvest-treated with calcium chloride and M. guilliermondii alone or in combination, respectively
2.4 CaCl 2不同处理时间对芒果采后炭疽病抑制效果的影响
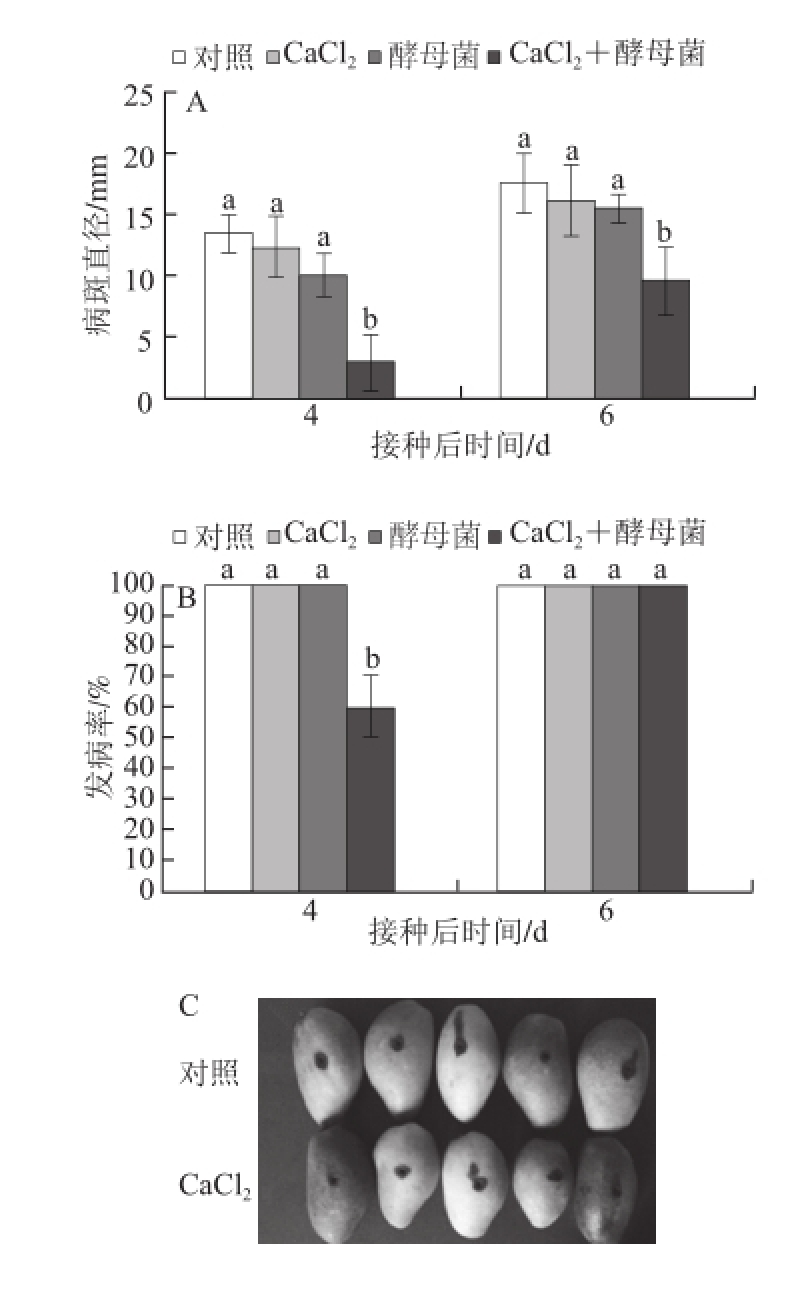

图4 CaaCCll
2不同处理时间对芒果采后炭疽病抑制效果的影响
Fig.4 Effect of different incubation times with calcium chloride on control of C. gloeosporioides in mango fruit
A.病斑直径;B.发病率;C. CaCl 2先于病原菌2 h接种果实伤口;D. CaCl 2先于病原菌24 h接种果实伤口。
由图4可知,CaCl 2不同时间处理对芒果采后炭疽病具有不同的抑制效果。接种间隔时间为24 h时,在28 ℃条件下贮藏4 d后,芒果采后炭疽病病斑直径和发病率仅为2.55 mm和60%,显著低于对照果实的13.54 mm和100%(图4A、B)。CaCl 2先于C. gloeosporioides孢子2 h对果实进行处理时,对芒果采后炭疽病的抑制效果与对照相比,无显著差异(图4C),而CaCl 2与C. gloeosporioides孢子处理间隔时间扩大到24 h时,对芒果采后炭疽病的抑制效果要显著好于对照(图4D)。
2.5 CaCl 2和拮抗菌M. guilliermondii处理对芒果果实PAL、POD和PPO活性的影响
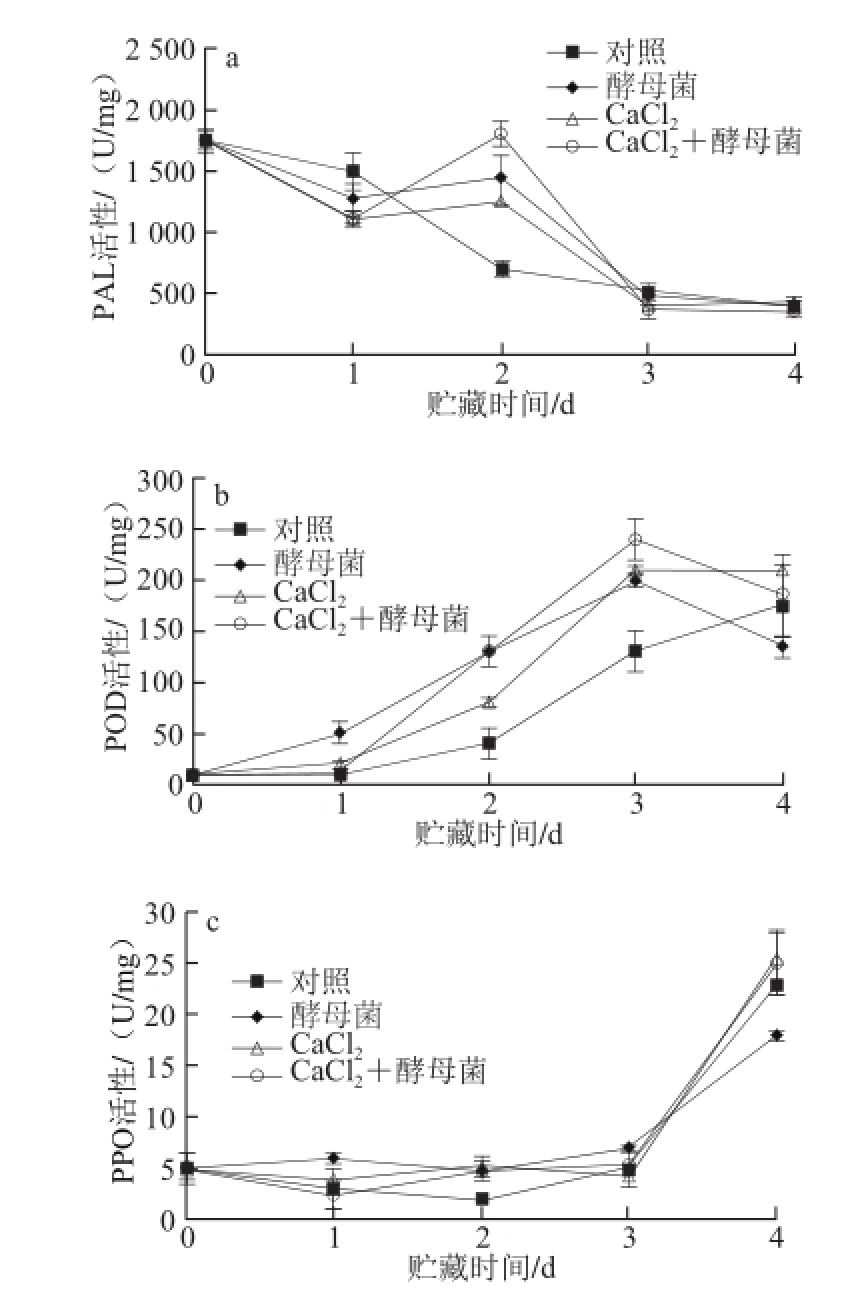
图5 CaCl
2和M. guilliermondii对采后芒果果实PAL(a)、POD(b)和PPO(c)活性的影响
Fig.5 Changes in activities of PAL (a), PPO (b) and POD (c) of mango fruit postharvest-treated with calcium chloride and M. guilliermondii alone or in combination, respectively
由图5可知,在贮藏前1~4 d期间,对照组果实PAL活性处于逐渐下降的趋势,而其他处理在贮藏第1~3天期间均显著诱导了果实中PAL活性的上升,并在贮藏后第2天时到达峰值,其中滴加40 g/L CaCl 210 min后接种拮抗菌M. guilliermondii处理的果实要显著高于其他处理,此时PAL活性是对照果实的2.45 倍(图5a)。在贮藏前3 d,各处理芒果果实POD活性始终保持上升的趋势,在贮藏第3天时到达峰值。其中,滴加40 g/L CaCl 2或单独接种拮抗菌M. guilliermondii处理的果实P O D活性都要显著高于对照,达到1.76 倍。同样,滴加40 g/L CaCl 210 min后接种拮抗菌M. guilliermondii处理的果实POD活性要显著高于单一处理的果实,随着芒果果实贮藏时间的延长,各处理POD活性都逐渐下降(图5b)。所有处理芒果果实PPO活性在贮藏前3 d基本保持稳定,但随后酶活性急剧升高。与对照相比,滴加40 g/L CaCl 210 min后接种拮抗菌M. guilliermondii处理的果实PPO活性显著提升(图5c)。
前期研究发现拮抗菌M. guilliermondii处理对芒果果实采后炭疽病具有一定的抑制作用,但同时也存在有效处理质量浓度偏高和生防效力低下等不足。因此,为进一步提高拮抗菌M. guilliermondii的生防效力,本实验测试了通过使用CaCl 2来提高拮抗菌M. guilliermondii拮抗效力的可行性。实验发现,质量浓度为5~20 g/L的CaCl 2对C. gloeosporioides的生长具有促进作用,但质量浓度过大(50 g/L)时具有抑制作用。表现在40 g/L CaCl 2能够提高拮抗菌M. guilliermondii在芒果果实伤口处的增殖速度,而50 g/L CaCl 2则抑制拮抗菌M. guilliermondii在芒果果实伤口处的生长。所以CaCl 2与拮抗菌M. guilliermondii的复合处理的最佳质量浓度为应为40 g/L。结果证实,40 g/L CaCl 2能显著提高拮抗菌M. guilliermondii对芒果采后炭疽病的生防效力,其中复合处理对芒果采后炭疽病的抑制效果较拮抗菌M. guilliermondii单一处理更为显著。
大量研究 [23-25]表明,抗病相关酶PAL、POD和PPO活性的变化与果蔬采后对各种病菌的防卫反应能力密切相关。之前有研究发现,采后果蔬利用生物和非生物诱导因子处理,都能够提高果蔬抗病相关酶的表达,增强果蔬对病害的抵抗能力 [26-28]。本实验结果显示,40 g/L CaCl 2与C. gloeosporioides孢子处理间隔时间扩大到24 h时,对芒果采后炭疽病的抑制效果显著增强,这说明CaCl 2能够诱导芒果果实产生抗病性。这一结果与前人的报道相类似,研究 [29]结果表明,CaCl 2处理的梨果实能够诱导其产生抗病性,从而增强梨对青霉病和灰霉病的防治效果。另外,通过对芒果果实抗病相关酶酶活性进行测定,结果同样表明,40 g/L CaCl 2和拮抗菌M. guilliermondii均可显著诱导芒果果实抗病相关酶PAL、POD和PPO活性的上升,其中复合处理对芒果果实抗病相关酶的诱导比单一处理更显著。
综上,40 g/L CaCl 2能够直接抑制炭疽病原菌的生长和促进拮抗菌M. guilliermondii在芒果果实伤口处的增殖,同时还具有诱导芒果果实产生抗病性的作用。因此,可综合提高拮抗菌M. guilliermondii生防效力,从而减少芒果实采后炭疽病病害的发生,在芒果采后的运输和贮藏过程中显示了良好的应用前景。
参考文献:
[1] 黄辉白. 热带亚热带果树栽培学[M]. 北京: 高等教育出版社, 2003: 106.
[2] DODD J C, PRUSKY D, JEFFRIES P. Fruit diseases[M]//LITZ R E. The mango: botany, production and uses. Cambridge: CAB International, 1997: 257-280.
[3] JOHNSON G I, SHARP J L, MILNE D L, et al. Postharvest technology and quarantine treatments[M]//LITZ R E. The mango: botany, production and uses. Cambridge: CAB International, 1997: 447-507.
[4] KOBILER I, SHALOM Y, ROTH I, et al. Effect of 2,4-dichlorophenoxyacetic acid on the incidence of side and stem end rots in mango fruits[J]. Postharvest Biology and Technology, 2001, 23(1): 23-32. DOI:10.1016/S0925-5214(01)00092-8.
[5] DROBY S, WISNIEWSKI M, MACARISIN D, et al. Twenty years of postharvest biocontrol research: is it time for a new paradigm?[J]. Postharvest Biology and Technology, 2009, 52: 137-145. DOI:10.1016/j.postharvbio.2008.11.009.
[6] JANISIEWICZ W J, CONWAY W S. Combining biological control with physical and chemical treatments to control fruit decay after harvest[J]. Stewart Postharvest Review, 2010, 6(1): 1-16. DOI:10.2212/spr.2010.1.3.
[7] SHARMA R R, SINGH D, SINGH R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: a review[J]. Biological Control, 2009, 50(3): 205-221. DOI:10.1016/ j.biocontrol.2009.05.001.
[8] LIU H M, GUO J H, LUO L, et al. Improvement of Hansenias porauvarum biocontrol activity against gray mold by the addition of ammonium molybdate and the possible meehanisms involved[J]. Crop Protection, 2010, 29: 277-282.
[9] YU T, LI H Y, ZHENG X D. Synergistic effect of chitosan and Cryptococcus laurentii on inhibition of Penicillium expansum infections[J]. International Journal of Food Microbiology, 2007, 114(3): 261-266. DOI:10.1016/j.ijfoodmicro.2006.09.008.
[10] QIN G Z, TIAN S P, XU Y, et al. Combination of antagonistic yeasts with two food additives for control of brown rot caused by Monilinia fructicola on sweet cherry fruit[J]. Journal of Applied Microbiology, 2005, 100(3): 508-515. DOI:10.1111/j.1365-2672.2005.02821.x.
[11] YAO H J, TIAN S P, WANG Y S . Sodium bicarbonate enhances biocontrol ef cacy of yeasts on fungal spoilage of pears[J]. International Journal of Food Microbiology, 2004, 93(3): 297-304.
[12] ZHANG H Y, FU C X, ZHENG X D, et al. Control of postharvest Rhizopus rot of peach by microwave treatment and yeast antagonist[J]. European Food Research and Technology, 2004, 218(6): 568-572. DOI:10.1007/s00217-004-0902-9.
[13] YU T, CHEN J S, CHEN R, et al. Biocontrol of blue and gray mold diseases of pear fruit by integration of antagonistic yeast with salicylic acid[J]. International Journal of Food Microbiology, 2007, 116(3): 339-345. DOI:10.1016/j.ijfoodmicro.2007.02.005.
[14] ZHANG H Y, ZHENG X D, WANG L, et al. Effect of yeast antagonist in combination with hot water dips on postharves Rhizopus rot of strawberries[J]. Journal of Food Engineering, 2007, 78: 281-287. DOI:10.1016/j.jfoodeng.2005.09.027.
[15] MARTIN-DIANA A B, RICO D, FRIAS J M, et al. Calcium for extending the shelf life of fresh whole and minimally processed fruits and vegetables: a review[J]. Trends in Food Science and Technology, 2007, 18(4): 210-218. DOI:10.1016/j.tifs.2006.11.027.
[16] WANG Y F. Control of postharvest decay on cherry tomatoes by marine yeast Rhodosporidium paludigenum and calcium chloride[J]. Journal of Applied Microbiology, 2010, 109(2): 651-656. DOI:10.1111/j.1365-2672.2010.04693.x.
[17] GHOLAMNEJAD J, ETEBARIAN H R. Effect of calcium chloride on the biocontrol efficacy of two antagonistic yeasts against Penicillium expansum on apple fruit[J]. Phytoparasitica, 2009, 37(3): 255-261. DOI:10.1007/s12600-009-0033-8.
[18] JANISIEWICZ W J, USALL J, BORS B. Nutritional enhancement of biocontrol of blue mold on apples[J]. Phytopathology, 1992, 82(11): 1364-1370.
[19] ASSIS J S, MALDONADO R, MUNOZ T, et al. Effect of high carbon dioxide concentration on PAL activity and phenolic contents in ripening cherimoya fruit[J]. Postharvest Biology and Technology, 2001, 23(1): 33-39. DOI:10.1016/S0925-5214(01)00100-4.
[20] CHANCE B, MAEHLY A C. Assay of catalases and peroxidases[J]. Methods in Enzymology, 2005, 2: 764-817.
[21] WANG Y S, TIAN S P, XU Y, et al. Changes in the activities of proand anti-oxidant enzymes in peach fruit inoculated with Cryptococcus laurentii or Penicillium expansum at 0 or 20 ℃[J]. Postharvest Biology and Technology, 2004, 34(1): 21-28. DOI:10.1016/ j.postharvbio.2004.04.003.
[22] BRADFORD M M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of proteindye binding[J]. Analytical Biochemistry, 1976, 72: 248-254.
[23] MOHAMMADI M, KAZEMI H. Changes in peroxidase and polyphenol oxidase activities in susceptible and resistant wheat heads inoculated with Fusarium graminearum and induced resistance[J]. Plant Science, 2002, 162(4): 491-498. DOI:10.1016/S0168-9452(01)00538-6.
[24] MILOSEVIC N, SLUSARENKO A J. Active oxygen metabolism and lignification in the hypersensitive response in bean[J]. Physiological and Molecular Plant Pathology, 1996, 49(3): 143-157. DOI:10.1006/ pmpp.1996.0045.
[25] PASSARDI F, PENEL C, DUNAND C. Performing the paradox-ical: how plant peroxidases modify the cell wall[J]. Trends in Plant Science, 2004, 9(11): 534-540. DOI:10.1016/j.tplants.2004.09.002.
[26] LI R P, ZHANG H Y, LIU W M, et al. Biocontrol of postharvest gray and blue mold decay of apples with Rhodotorula mucilaginosa and possible mechanisms of action[J]. International Journal of Food Microbiology, 2011, 146: 151-156. DOI:10.1016/ j.ijfoodmicro.2011.02.015.
[27] HERSHKOVITZ V, BEN-DAYAN C, RAPHAEL G, et al. Global changes in gene expression of grapefruit peel tissue in response to the yeast biocontrol agent Metschnikowia fructicola[J]. Molecular Plant Pathology, 2012, 13(4): 338-349.
[28] ZHENG X L, YE L B, JIANG T J, et al. Limiting the deterioration of mango fruit during storage at room temperature by oxalate treatment[J]. Food Chemistry, 2012, 130: 279-285. DOI:10.1016/ j.foodchem.2011.07.035.
[29] YU T, YU C, ZHENG X D. Effect of Cryptococcus laurentii and calcium chloride on control of Penicillium expansum and Botrytis cinerea infections in pear fruit[J]. Biological Control, 2012, 61: 169-175. DOI:10.1016/j.biocontrol.2012.01.012.
Control of Postharvest Anthracnose of Mango Fruit by Combining Yeast Meyerozyma guilliermondii with Calcium Chloride
CHEN Min
1, GAO Yunkai
1, SONG Haichao
1, HE Shuting
1, ZHANG Rongyi
1, ZHONG Liwen
2, SHI Xuequn
3,*
(1. College of Environment and Plant Protection, Hainan University, Haikou 570228, China; 2. College of Horticulture and Landscape, Hainan University, Haikou 570228, China; 3. College of Food Science and Technology, Hainan University, Haikou 570228, China)
Abstract:The purposes of the present study were to investigate the effect of the combined treatment with calcium chloride and the antagonistic yeast Meyerozyma guilliermondii for control of postharvest athracnose caused by Colletotrichum gloeosporioides on mango fruit and to analyze the effect of CaCl 2on the growth of the pathogen and yeast as well as the disease resistance. The results indicated that the biocontrol activity of M. guilliermondii at 1 × 10 8CFU/mL on anthracnose decay was enhanced by co mbination with 40 g/L CaCl 2treatment. The combi ned treatment with M. guilliermondii and 40 g/L CaCl 2resulted in a remarka bly improved control of the disease in comparison with the treatment with M. guilliermondii or 40 g/L CaCl 2alone. In addition, 40 g/L CaCl 2inhibited mycelial growth of pathogens and enhanced the population of M. guilliermondii in the wounds of mango fruits. Meanwhile, the combined treatment induced higher defense-related enzymes act ivities than either alone. Thus, these results suggested that 40 g/L CaCl 2could improve the bio control activity of M. guilliermondii on the control of postharvest athracnose deca y caused by C. gloeosporioides in mango fruit, directly by inhibiting mycelial growth of pathogens and increasing population size of antagonist and indire ctly by enhancing disease resistance.
Key words:mango; Colletotri chum gloeosporioides; calcium chloride; Meyerozyma guilliermondii; bio control; enzyme activity
DOI:10.7506/spkx1002-6630-201602036
中图分类号:Q939.92;TS255.3
文献标志码:A
文章编号:1002-6630(2016)02-0204-06
引文格式:
陈敏, 高云慨, 宋海超, 等. 氯化钙结合季也蒙毕赤酵母(Meyerozyma guilliermondii)对抑制芒果采后炭疽病效果的影响[J].食品科学, 2016, 37(2): 204-209. DOI:10.7506/spkx1002-6630-201602036. http://www.spkx.net.cn
CHEN Min, GAO Yunkai, SONG Haichao, et al. Control of postharvest anthracnose of mango fruit by combining yeast Meyerozyma guilliermondii with calc ium chloride[J]. Food Science, 2016, 37(2): 204-209. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201602036. http://www.spkx.net.cn
收稿日期:2015-01-25
基金项目:国家自然科学基金面上项目(31360415);海南省自然科学基金项目(312061);海南大学中西部创新项目(MWECSP-RT08;ZXBJH-XK004;ZXBJH-XK005);海南省教育厅科研重点项目(HNKY2014-01)
作者简介:陈敏(1992—),女,硕士研究生,研究方向为热带水果采后病理学。E-mail:18805140991@163.com
*通信作者:史学群(1972—),男,副教授,博士,研究方向为采后病理学。E-mail:shixuequn@163.com