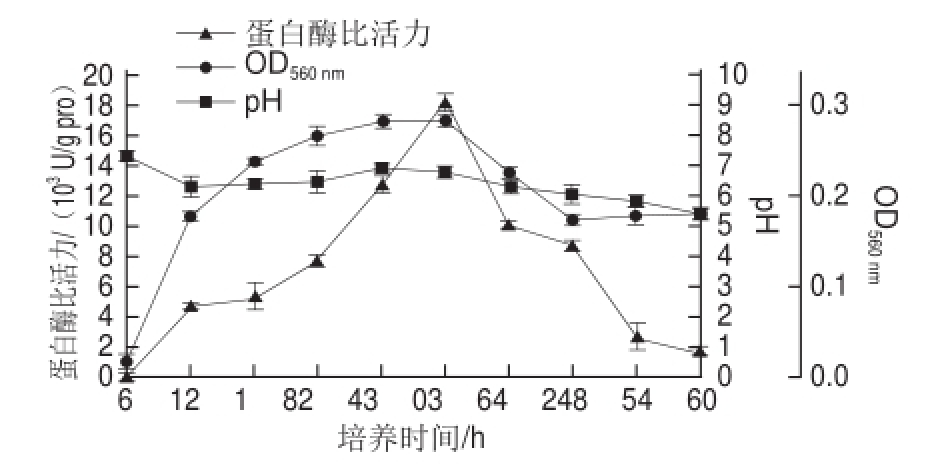
图1 蛋白酶发酵动力学曲线
Fig.1 Kinetic curves of protease activity and pH during fermentation
王 颖 1,高 翔 2,周 君 1,张春丹 1,李 晔 1,王祖忠 1,袁 贝 1,戴 娟 1,钱琴莲 1,苏秀榕 1, *
(1.宁波大学海洋学院,浙江 宁波 315211;2.浙江省乔司监狱,浙江 杭州 310019)
摘 要:为了优化地衣芽孢杆菌(Bacillus licheniformis)以浒苔(Enteromorpha prolifera)为碳源进行发酵产蛋白酶的能力,以蛋白酶比活力为指标,通过单因素试验探讨不同因素对产酶效率的影响,在此基础上运用Plackett-Burman设计筛选出了3 个显著因素:培养基初始pH值、浒苔添加量和NaH 2PO 4添加量。采用响应面法对这3 个显著因素进行优化,得到最佳产酶条件为培养基初始pH 6.66、浒苔添加量4.65%、NaH 2PO 4添加量0.027 4%,此条件下蛋白酶比活力预测值为66 354.70 U/g pro,3 次实验验证值为66 966.37 U/g pro。验证值与基础发酵条件下的最大蛋白酶比活力18 206.16 U/g pro相比,提高了2.68 倍。
关键词:地衣芽孢杆菌;浒苔;发酵条件;蛋白酶;Plackett-Burman设计
近年来,随着全球气候变化和水体富营养化,赤潮、绿潮等频频暴发,严重影响了海洋及周边的生态环境,威胁了养殖业和旅游业的发展 [1]。浒苔(Enteromorpha prolifera)等绿潮在我国青岛海域连续暴发,成为了全世界的研究热点 [2-5]。浒苔是一种繁殖迅速的大型绿藻,营养丰富,高膳食纤维、低脂肪,且富含维生素、矿物质等,浒苔干物质中碳水化合物含量为43.3%~60.2%,蛋白质含量为16.0%~22.1%,灰分含量为12.4%~18.7% [6]。碳水化合物是浒苔的主要成分 [6],利用浒苔作为发酵碳源,变废为宝,是浒苔资源化利用的一个理想途径。
蛋白酶是一种重要的工业酶制剂,在改善食品风味、提高食品品质等方面发挥着重要作用,是食品工业中应用最广泛的酶 [7-8]。本实验室从经 60Co辐照的东海香参中分离到一株能够产碱性蛋白酶的地衣芽孢杆菌(Bacillus licheniformis) [9]。与中性蛋白酶相比,碱性蛋白酶具有更强的水解能力和耐碱能力,有较强耐热性且有一定的酯酶活力,更易于实现工业化生产 [10]。地衣芽孢杆菌是目前碱性蛋白酶的主要生产菌株之一 [11],优化其发酵条件,提高其产酶能力,对工业扩大化生产具有重要意义。
本实验以地衣芽孢杆菌为发酵菌株,研究不同因素对发酵产酶能力的影响,采用Plackett-Burman试验设计筛选对发酵产酶有显著效应的因素,以浒苔为碳源,运用响应面分析法(response surface method,RSM)优化发酵条件,旨在为地衣芽孢杆菌产蛋白酶工业扩大化生产和浒苔的资源化利用提供理论依据。
1.1 材料与试剂
浒苔 福建福清市海兴保健食品有限公司。
Folin-酚试剂、酪氨酸标准品 美国Sigma公司;干酪素 北京市海淀区微生物培养基制品厂;其他化学试剂均为分析纯。
1.2 菌种与培养基
地衣芽孢杆菌(B. licheniformis)CGMCC 4323宁波市健康食品与海洋药物重点实验室分离、保藏。
种子斜面培养基:蛋白胨1.0%、牛肉膏0.3%、NaCl 0.5%、琼脂粉2.0%,自然pH值。种子液体培养基中不加琼脂粉。
基础发酵培养基:蔗糖7.500%、干酪素3.000%、NaCl 0.500%、K 2HPO 4·3H 2O 0.530%、NaH 2PO 4·2H 2O 0.030%、Na 2CO 30.056%、MnSO 40.002%,自然pH值。
1.3 方法
1.3.1 培养方法
种子培养:将-80 ℃保藏菌种划线接入种子斜面培养基,37 ℃静置培养12 h后接一环于种子液体培养基,37 ℃、120 r/min摇瓶培养12 h。
发酵培养:按6%接种量接种液体种子于本实验设置的不同发酵培养基中,37 ℃、120 r/min发酵培养36 h。
1.3.2 发酵条件的优化
在基础发酵培养基水平上分别对碳源、氮源、金属离子、初始pH值、接种量进行单因素优化。在单因素试验基础上进行试验次数为12 次的Plackett-Burman筛选试验,对浒苔添加量、酵母膏添加量、K 2HPO 4添加量、Na 2CO 3添加量、FeCl 3添加量、NaH 2PO 4添加量、培养基初始pH值、接种量8 个因素进行研究,筛选出有显著效应的关键因素。采用Box-Behnken原理设计对Plackett-Burman试验筛选出的3 个关键因素:培养基初始pH值、浒苔添加量和NaH 2PO 4添加量进行响应面优化,以获得较好的发酵条件。
1.3.3 分析方法
采用Folin-酚显色法测定蛋白酶活力。蛋白酶活力单位定义为:在40 ℃、pH 10.0的条件下,1 min内水解酪蛋白产生1 μg酪氨酸所需要的酶量为一个酶活力单位(U)。在本研究中,蛋白酶活力以酶比活力(U/g pro)表示。
菌体计数采用比浊计数法,以培养基为空白,测定OD 560 nm值。OD 560 nm值代表了菌体生物量,值越高表示菌体生物量越大。
1.4 数据处理
采用Design-Expert 8.0.6.1和SPSS 13.0进行数据分析,Origin 8.0软件制图。
2.1 基础发酵参数测定
在基础发酵培养基水平上,接种6%液体种子,每6 h取样,测定OD 560 nm、pH值和酶比活力,结果见图1。种子发酵6 h后菌体生长进入对数期,30 h后进入稳定期,36 h后菌体开始衰亡自溶,进入衰亡期。发酵起始12 h,培养基pH值急剧下降到6.2,之后慢慢回升到6.7左右,36 h后pH值又开始下降。乳酸是地衣芽孢杆菌的主要代谢产物 [12],pH值在菌体生长对数期急剧下降,可能与有机酸的产生有关;之后回升可能是由于菌体的代谢途径发生改变,有机酸在某些代谢途径中被利用或者菌体自身生长调节机制造成的。前6 h不产蛋白酶,24 h后大量产酶且在36 h时达到最大值,酶比活力为18 206.16 U/g pro,之后随着菌体的衰亡,pH值的下降,酶比活力也急剧下降。根据细胞生长与产酶的关系,可以把酶的生物合成模式分为同步合成、延续合成、中期合成和滞后合成4 种类型 [13]。分析图1中酶活力与细菌生长曲线发现,在地衣芽孢杆菌生长一段时间后碱性蛋白酶开始进行生物合成,并且当菌体生长进入稳定期后该酶还可以延续合成一段时间,因此可以推测地衣芽孢杆菌产碱性蛋白酶的生物合成属于延续合成型,产物形成属于部分生长偶联型,这一结果与马永强等 [13]研究地衣芽孢杆菌2709产碱性蛋白酶发酵动力学获得的结论一致。
图1 蛋白酶发酵动力学曲线
Fig.1 Kinetic curves of protease activity and pH during fermentation
2.2 单因素试验优化结果
2.2.1 碳源添加量对发酵产酶的影响
在基础发酵培养基水平上,以浒苔代替蔗糖作为碳源,浒苔添加量分别为2.5%、3.5%、4.5%、5.5%、6.5%、7.5%、8.5%、9.5%。浒苔添加量为4.5%时酶比活力最高,为53 469.64 U/g pro,与基础发酵最大酶比活力18 206.16 U/g pro相比提高了1.94 倍(图2),由此可见,浒苔比蔗糖更有利于地衣芽孢杆菌产蛋白酶。地衣芽孢杆菌产碱性蛋白酶的生物合成属于延续合成型,这种生物合成类型能够受到诱导物的诱导作用,而浒苔干物质中蛋白质的含量较高(16.00%~22.10%) [5],因此浒苔可能对蛋白酶的合成起到了诱导作用。同时,浒苔中纤维素(13.30%)与半纤维素(28.01%)的含量较高 [14],推测该菌株地衣芽孢杆菌主要以浒苔中的纤维素、半纤维素物质作为碳源进行发酵产酶,其胞外酶可能含纤维素酶,但其酶活性、降解纤维素的能力及浒苔作为碳源的利用效率还有待研究。另外,浒苔中氨基酸含量丰富,为13.30%(以干样计),且种类齐全,主要包括天冬氨酸(1.52%)、苯丙氨酸(0.71%)、赖氨酸(0.68%)、亮氨酸(1.22%)、异亮氨酸(0.58%)、苏氨酸(0.75%)、丝氨酸(0.74%)、组氨酸(0.22%)等 [15]。这些氨基酸对地衣芽孢杆菌的生长有明显促进作用,尤其是赖氨酸,不仅有利于生长,而且具有特殊的刺激生长作用,可视为生长因子 [12],这也可能是浒苔作为碳源能够大大提高地衣芽孢杆菌产酶的原因之一。
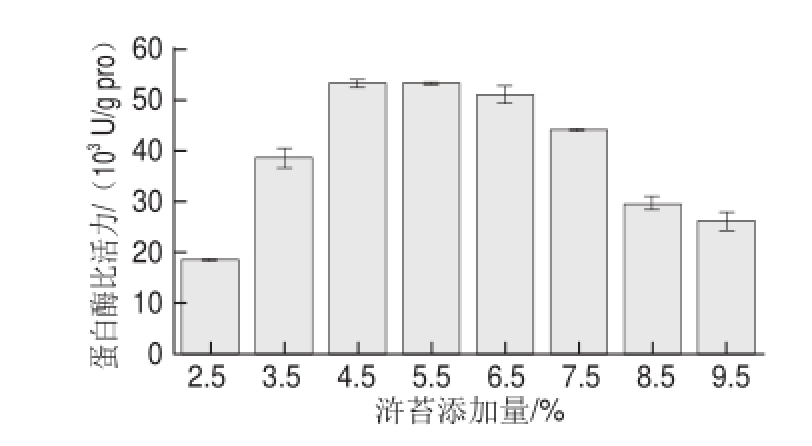
图2 浒苔添加量对蛋白酶活力的影响
Fig.2 Effect of E. prolifera concentration on protease activity
2.2.2 氮源对发酵产酶的影响
在基础发酵培养基水平上,分别添加3%的干酪素、牛肉膏、酵母膏、大豆蛋白、明胶、尿素、硝酸铵和蛋白胨作为氮源。图3a表明,不同氮源对产酶影响有较大差异,本实验中有机氮源比无机氮源更利于发酵产酶,尤其添加酵母膏作为氮源时蛋白酶比活力最高,为48 583.78 U/g pro,其次是大豆分离蛋白、明胶和尿素,添加硝酸铵和蛋白胨时蛋白酶比活力为0 U/g pro。孙倩等 [16]对地衣芽孢杆菌产碱性蛋白酶的研究表明,以豆粕和磷酸铵混合物作为氮源时酶活最高,有机氮源与无机氮源的组合可以更充分地为菌种提供营养物质。本研究仅对单一氮源的选择进行了初步筛选,不同氮源组合的筛选有待研究。
进一步研究酵母膏添加量对发酵产酶的影响,酵母膏添加量分别为1%、2%、3%、4%、5%、6%、7%、8%。酵母膏添加量为4%时酶比活力最高,为57 699.15 U/g pro,与基础发酵最大酶比活力18 206.16 U/g pro相比提高了2.17 倍(图3b)。
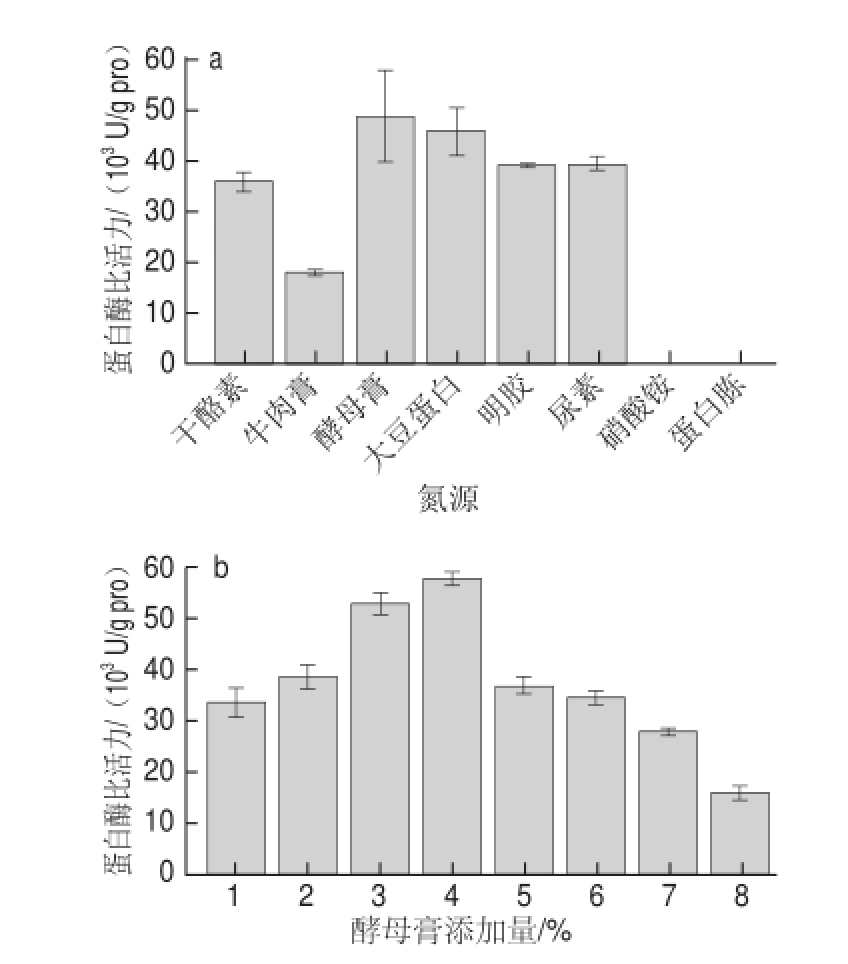
图3 不同氮源及酵母膏添加量对蛋白酶活力的影响
Fig.3 Effect of nitrogen sources and yeast extract concentration on protease activity
2.2.3 金属离子对发酵产酶的影响
在基础发酵培养基水平上,添加0.002%不同金属离子。由图4可知,添加Fe 3+时酶比活力最高,为55 007.92 U/g pro,与基础发酵最大酶比活力18 206.16 U/g pro相比提高了2.02 倍,其次是Li +和Ba 2+,添加Mg 2+酶比活力最低。这表明Fe 3+对地衣芽孢杆菌产蛋白酶有促进作用。

图4 不同金属离子对蛋白酶活力的影响
Fig.4 Effect of different metal ions on protease activity
2.2.4 初始pH值对发酵产酶的影响
在基础发酵培养基水平上,调节培养基初始pH值为5、6、7、8、9、10、11、12。由图5可知,初始pH值为7时酶比活力最高,为58 651.32 U/g pro,与基础发酵最大酶比活力18 206.16 U/g pro相比提高了2.22 倍。pH值继续增大时,酶比活力逐渐降低,说明在中性培养基条件下地衣芽孢杆菌产蛋白酶活力较高。这一结果与刘海进 [17]、卜令军 [18]等在地衣芽孢杆菌产碱性蛋白酶培养条件优化中确定的初始pH值为6.5~7.5一致,说明中性培养基条件下能够稳定维持菌体生长与产酶效率。
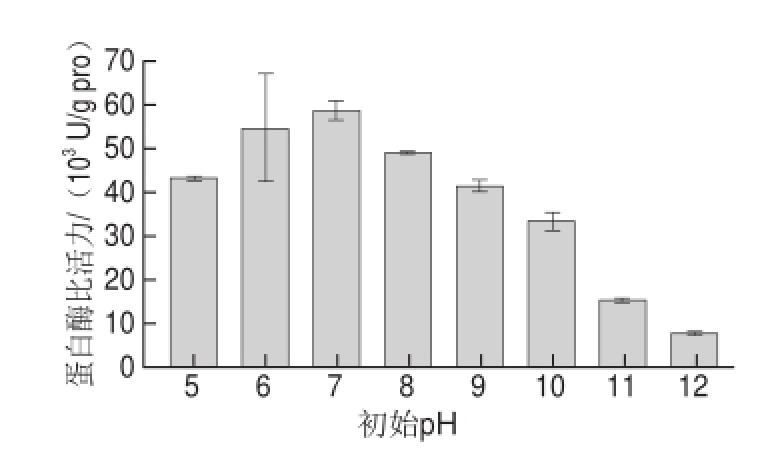
图5 初始pH值对蛋白酶活力的影响
Fig.5 Effect of initial medium pH on protease activity
2.2.5 接种量对发酵产酶的影响
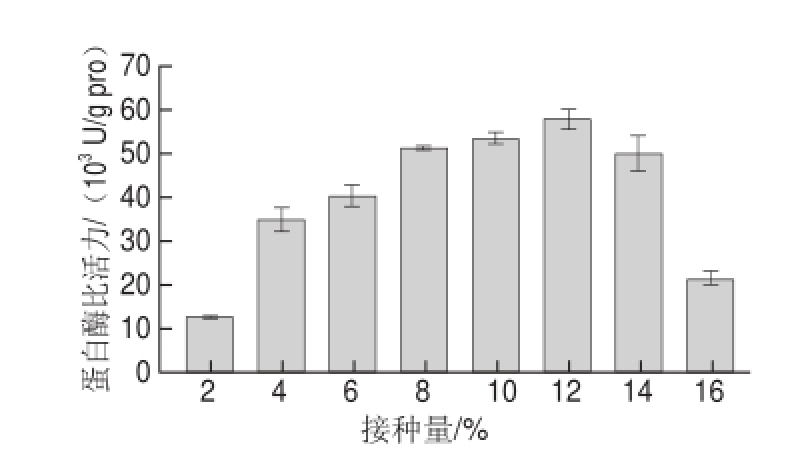
图6 接种量对蛋白酶活力的影响
Fig.6 Effect of inoculum amount on protease activity
分别接种2%、4%、6%、8%、10%、12%、14%、16%的培养12 h种子培养基于基础发酵培养基中。由图6可知,接种量为12%时酶比活力最大,为57 854.18 U/g pro,与基础发酵最大酶比活力18 206.16 U/g pro相比提高了2.18 倍。随后蛋白酶比活力呈现下降趋势,可能是因为接种量增大,菌体进入对数期时间较早,培养基中菌体含量过多,导致营养成分不能满足菌体正常生长代谢,从而影响了产蛋白酶能力。
2.3 Plackett-Burman筛选试验
Plackett-Burman试验设计及响应值见表1。从显著性检验结果(表2)可以看出,各影响因素的显著性排序为:初始pH值>浒苔添加量>NaH 2PO 4添加量>K 2HPO 4添加量>FeCl 3添加量>接种量>酵母膏添加量>Na 2CO 3添加量。其中,初始pH值、浒苔添加量和NaH 2PO 4添加量对蛋白酶比活力的影响最为显著。因此,选取这3 个关键因素进一步作响应面分析。其他因素根据单因素试验和节约成本的原则,将其水平控制在较好水平。
表1 Plackett-Burman试验设计与结果(n=8)
Table 1 Plackett-Burman design with experimental values of protease activity (n= 8)

试验号 A浒苔添加量/% Y酶比活力/(U/g pro)1 7.0 6 0.5 0.10 0.02 0.06 7 6 71 083.18 2 3.5 6 1.0 0.05 0.02 0.06 9 6 8 719.10 3 7.0 3 1.0 0.10 0.01 0.06 9 12 34 689.62 4 3.5 6 0.5 0.10 0.02 0.03 9 12 16 478.20 5 3.5 3 1.0 0.05 0.02 0.06 7 12 53 328.57 6 3.5 3 0.5 0.10 0.01 0.06 9 6 20 237.91 7 7.0 3 0.5 0.05 0.02 0.03 9 12 45 379.35 8 7.0 6 0.5 0.05 0.01 0.06 7 12 63 656.55 9 7.0 6 1.0 0.05 0.01 0.03 9 6 4 422.73 10 3.5 6 1.0 0.10 0.01 0.03 7 12 10 040.90 11 7.0 3 1.0 0.10 0.02 0.03 7 6 40 200.80 12 3.5 3 0.5 0.05 0.01 0.03 7 6 21 581.45 B酵母膏添加量/% C K 2HPO 4添加量/% D Na 2CO 3添加量/% E FeCl 3添加量/% F NaH 2PO 4添加量/% G初始pH H接种量/%
表2 回归方程显著性检验
Table 2 Significance test of regression equation

因素 自由度 参数估计 F值 P值 显著性排序A 1 10 753.842 0 84.356 1 0.002 7 2 B 1 -3 418.086 3 8.522 3 0.061 5 7 C 1 -7 251.241 9 38.354 3 0.008 5 4 D 1 -363.093 7 0.096 2 0.776 8 8 E 1 6 713.336 9 32.875 0 0.010 5 5 F 1 9 467.624 1 65.384 0 0.004 0 3 G 1 -10 830.380 0 85.561 2 0.002 7 1 H 1 4 777.334 8 16.648 0 0.026 6 6
2.4 响应面法优化发酵条件
利用Design-Expert 8.0.6.1软件对表3数据进行二次回归分析,得到二次多元回归模型为:
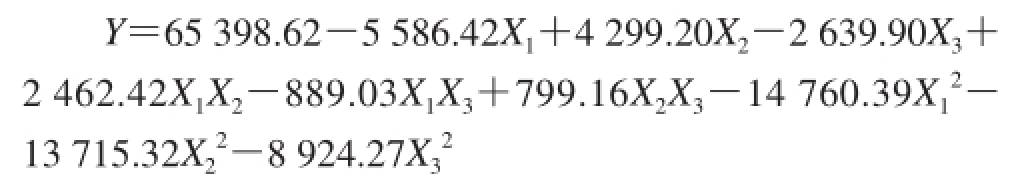
该方程表达了蛋白酶比活力(Y)与3 个自变量间X 1、X 2、X 3的关系。回归方程的一次项和平方项系数都较大,说明响应值与试验因子之间并不是简单的线性关系;而交互项系数较小,说明响应面分析所选3 个因素间的交互效应较小。变量的正系数表明该变量的正向变化可引起响应值的增加;负的二次项系数表明方程的抛物面开口向下,具有极大值点,能够进行最优分析。
表3 响应面试验设计方案与结果
Table 3 RSM Design with experimental values of protease activity
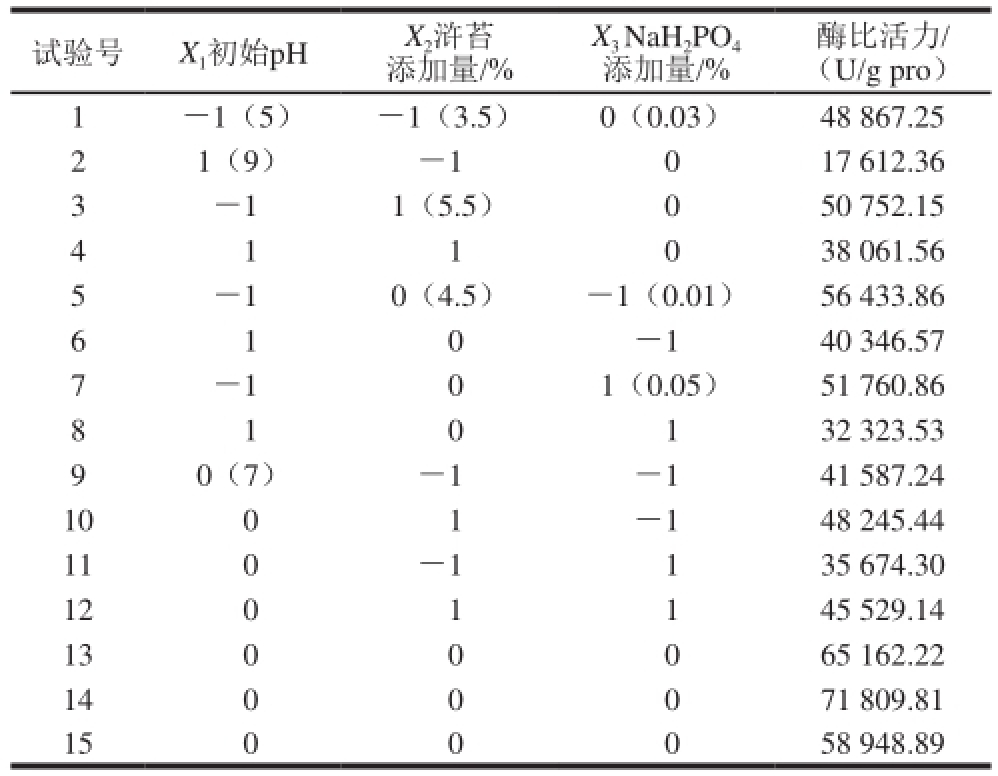
试验号 X 1初始pH X 2浒苔添加量/%酶比活力/(U/g pro)1 -1(5) -1(3.5) 0(0.03) 48 867.25 2 1(9) -1 0 17 612.36 3 -1 1(5.5) 0 50 752.15 4 1 1 0 38 061.56 5 -1 0(4.5) -1(0.01) 56 433.86 6 1 0-1 40 346.57 7 -1 0 1(0.05) 51 760.86 8 1 0 1 32 323.53 X 3NaH 2PO 4添加量/% 9 0(7) -1 -1 41 587.24 10 0 1 -1 48 245.44 11 0 -1 1 35 674.30 12 0 1 1 45 529.14 13 0 0 0 65 162.22 14 0 0 0 71 809.81 15 0 0 0 58 948.89
表4 响应面试验结果方差分析
Table 4 Analysis of variance of response surface regression model
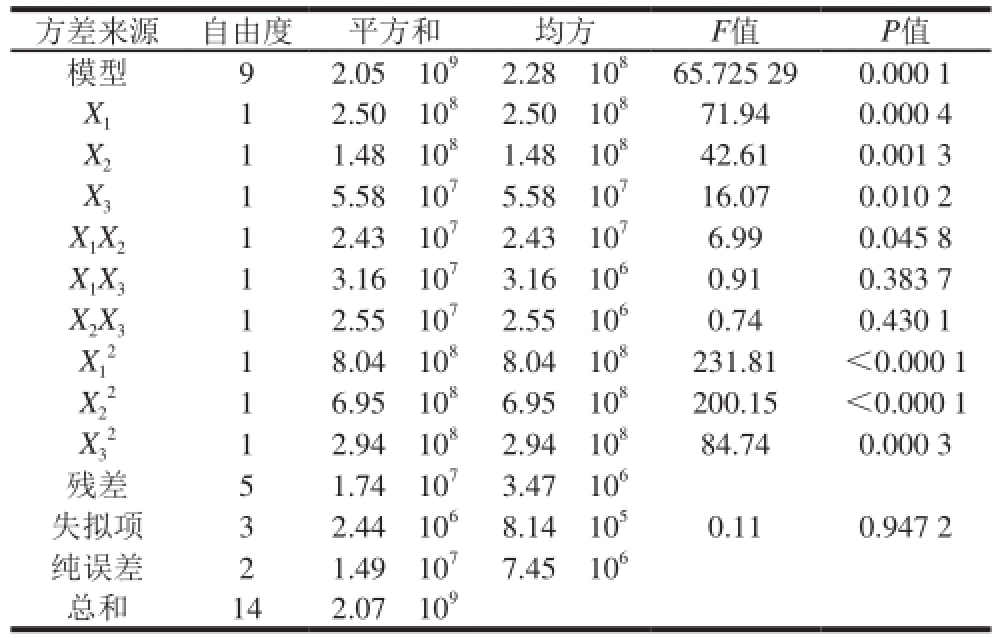
方差来源 自由度 平方和 均方 F值 P值模型 9 2.05h10 92.28h10 865.725 29 0.000 1 X 11 2.50h10 82.50h10 871.94 0.000 4 X 21 1.48h10 81.48h10 842.61 0.001 3 X 31 5.58h10 75.58h10 716.07 0.010 2 X 1X 21 2.43h10 72.43h10 76.99 0.045 8 X 1X 31 3.16h10 73.16h10 60.91 0.383 7 X 2X 31 2.55h10 72.55h10 60.74 0.430 1 X 11 8.04h10 88.04h10 8231.81 <0.000 1 X2 2 1 2.94h10 82.94h10 884.74 0.000 3残差 5 1.74h10 73.47h10 6失拟项 3 2.44h10 68.14h10 50.11 0.947 2纯误差 2 1.49h10 77.45h10 6总和 14 2.07h10 91 6.95h10 86.95h10 8200.15 <0.000 1 X3 22
回归模型的方差分析见表4。决定系数R 2为0.991 6,模型的P值小于0.01,说明该模型回归方程极显著,不同的试验因子之间差异高度显著,该试验方法是可靠的。失拟项的P值为0.947 2>0.05,差异不显著,表明回归模型拟合程度较好,可用于试验分析和预测。
响应面三维图是回归方程的图形表述,可从图中直观快速地找到最佳参数、参数之间的相互作用以及最大的响应值 [19],从图7上可以形象地看出各因素交互作用对响应值的影响。随着初始pH值的提高、浒苔添加量的增加、NaH 2PO 4添加量的增加,蛋白酶比活力都呈现出先升后降的趋势。响应面曲面坡度陡峭,表明各因素对酶活力的影响较大;等高线呈现椭圆型,各图的两因素交互作用明显。结果表明在中心点附近存在最大响应值。

图7 初始pH值、浒苔添加量、NaH
2PO
4添加量3 个因素间交互作用的响应面图
Fig.7 Response surface plots for the effects of initial medium pH,Enteromorpha prolifera concentration and NaH
2PO
4concentration on protease activity
对二次回归方程求解,当响应值Y最大时各因素的水平分别为X 1=-0.17、X 2=0.14、X 3=-0.13,转换后得最佳发酵条件为初始pH 6.66、浒苔添加量4.65%、NaH 2PO 4添加量0.027 4%,此条件下的理论预测蛋白酶比活力为66 354.70 U/g pro。为检验实验的可靠性,按照最优条件进行3 次实验验证,实际测得的蛋白酶比活力平均值为66 966.37 U/g pro,与理论预测值相比,相对误差为0.9%。说明采用响应面法优化得到的发酵条件参数准确可靠,具有实用价值。
本研究通过单因素试验、Plackett-Burman设计和响应面法相结合,确定了地衣芽孢杆菌发酵浒苔产碱性蛋白酶的最佳条件为初始pH 6.66、浒苔添加量4.65%、NaH 2PO 4添加量0.027 4%,此条件下的蛋白酶比活力预测值为 66 354.70 U/g pro,3 次实验验证平均值为 66 966.37 U/g pro,验证值与预测值基本相符,且与基础发酵最大蛋白酶比活力18 206.16 U/g pro相比,验证值提高了2.68 倍,地衣芽孢杆菌的产酶能力得到了显著提高。
很多研究表明,浒苔含有生物活性物质,具有显著的药理活性,如降血脂 [20-21]、抗氧化 [21-22]、提高免疫力 [22-24]和抑制皮肤癌 [25]等。本研究在获得蛋白酶的同时还获得了主要成分为浒苔残渣和地衣芽孢杆菌的发酵副产物,若能够对副产物的具体成分进行分析并充分发掘其生理功能,寻求提高其高值综合利用途径,将大大提高经济效益,并更好地服务于创建资源节约型社会,因此,发酵副产物的成分及生理活性功能也值得今后进一步探讨研究。
参考文献:
[1] 任厚朋. 浒苔的开发及综合应用[J]. 上海化工, 2014, 39(3): 1-4. DOI:10.3969/j.issn.1004-017X.2014.03.001.
[2] 孙伟红, 冷凯良, 王志杰, 等. 浒苔的氨基酸和脂肪酸组成研究[J]. 渔业科学进展, 2009, 30(2): 106-109. DOI:10.3969/ j.issn.1000-7075.2009.02.018.
[3] 罗佳捷, 肖淑华, 张彬, 等. 浒苔的研究进展及其在动物生产中的应用[J]. 饲料博览, 2014(2): 31-34.
[4] 裘迪红, 宋绍华, 苏秀榕, 等. 浒苔加热过程中挥发性风味成分的变化[J]. 中国食品学报, 2013, 13(5): 250-257.
[5] 孙元芹, 李翘楚, 李红艳, 等. 浒苔生理活性与开发利用研究进展[J]. 水产科学, 2013, 32(4): 244-248. DOI:10.3969/ j.issn.1003-1111.2013.04.013.
[6] LI Yinping, WANG Jin, YU Yuan, et al. Production of enzymes by Alteromonas sp. A321 to degrade polysaccharides from Enteromorpha prolifera[J]. Carbohydrate Polymers, 2013, 98(1): 988-994. DOI:10.1016/j.carbpol.2013.07.016.
[7] 何林玲, 何贝, 张霞. 蛋白酶在食品加工中的应用进展研究[J]. 食品工程, 2014(1): 12-14. DOI:10.3969/j.issn.1673-6044.2014.01.004.
[8] 胡学智, 王俊. 蛋白酶生产和应用的进展[J]. 工业微生物, 2008(4): 49-61. DOI:10.3969/j.issn.1001-6678.2008.04.010.
[9] 张文杰, 高翔, 孙静, 等. 地衣芽孢杆菌A产碱性蛋白酶的研究[J]. 食品科学, 2011, 32(1): 199-202.
[10] 包巨南, 兰立新, 肖怀秋. 微生物源碱性蛋白酶研究进展[J]. 酿酒, 2007, 34(3): 50-52. DOI:10.3969/j.issn.1002-8110.2007.03.020.
[11] 邓菊云. 微生物碱性蛋白酶研究进展[J]. 现代食品科技, 2008, 24(3): 293-296. DOI:10.3969/j.issn.1673-9078.2008.03.028.
[12] 胡尚勤, 刘天贵. 地衣芽孢杆菌营养要求的研究[J]. 河北省科学院学报, 2000, 17(4): 224-227. DOI:10.3969/ j.issn.1001-9383.2000.04.008.
[13] 马永强, 尹永智, 杨春华, 等. 地衣芽孢杆菌2709产碱性蛋白酶的酵动力学研究[J]. 食品工业科技, 2010, 31(6): 159-161; 164.
[14] 费岚, 邵飞, 贾睿, 等. 浒苔纤维素分解菌株的筛选、鉴定与纤维素酶活特性研究[J]. 上海海洋大学学报, 2014, 23(3): 411-416.
[15] ANTONIO G, 张威, 苏秀榕, 等. 浒苔和龙须菜营养成分的研究[J]. 水产科学, 2010, 29(6): 329-333. DOI:10.3969/ j.issn.1003-1111.2010.06.005.
[16] 孙倩, 陈复生, 丁长河, 等. 地衣芽孢杆菌产碱性蛋白酶发酵条件优化[J]. 食品工业科技, 2012, 33(13): 174-177; 192.
[17] 刘海进, 钱坤, 李吕木, 等. 地衣芽孢杆菌D-1产碱性蛋白酶培养条件的优化[J]. 激光生物学报, 2011, 20(3): 387; 409-412. DOI:10.3969/j.issn.1007-7146.2011.03.025.
[18] 卜令军, 李玉, 路福平, 等. 利用木糖母液生产碱性蛋白酶发酵工艺的初步研究[J]. 天津科技大学学报, 2014(6): 32-35; 40. DOI:10.13364/j.issn.1672-6510.2014.06.007.
[19] 孙静, 李晔, 张春丹, 等. 响应面法优化双酶水解鲣鱼下脚料[J]. 中国食品学报, 2013, 13(6): 104-110.
[20] TENG Zhaolin, QIAN Li, ZHOU Yan. Hypolipidemic activity of the polysaccharides from Enteromorpha prolifera[J]. International Journal of Biological Macromolecules, 2013, 62: 254-256. DOI:10.1016/ j.ijbiomac.2013.09.010.
[21] TANG Zhihong, GAO Hongwei, WANG Shuai, et al. Hypolipidemic and antioxidant properties of a polysaccharide fraction from Enteromorpha prolifera[J]. International Journal of Biological Macromolecules, 2013, 58: 186-189. DOI:10.1016/ j.ijbiomac.2013.03.048.
[22] ZHANG Zhongshan, WANG Xiaomei, ZHAO Mingxing, et al. The immunological and antioxidant activities of polysaccharides extracted from Enteromorpha linza[J]. International Journal of Biological Macromolecules, 2013, 57: 45-49. DOI:10.1016/ j.ijbiomac.2013.03.006.
[23] ROSARIO C, IGNACIO Z, JESUS L. Water-soluble seaweed extracts modulate the respiratory burst activity of turbot phagocytes[J]. Aquaculture, 2004, 229(Suppl 1/4): 67-78. DOI:10.1016/S0044-8486(03)00401-0.
[24] 徐大伦, 黄晓春, 杨文鸽, 等. 浒苔多糖的分离纯化及其对非特异性免疫功能的体外实验研究[J]. 中国食品学报, 2006, 6(5): 17-21. DOI:10.3969/j.issn.1009-7848.2006.05.004.
[25] HIQASHI-OKAJ K, OTANI S, OKAI Y. Potent suppressive effect of a Japanese edible seaweed, Enteromorpha prolifera (Sujiao-nori) on initiation and promotion phases of chemically induced mouse skin tumorigenesis[J]. Cancer Letters, 1999, 140(1/2): 21-25. DOI:10.1016/ S0304-3835(99)00304-3.
Optimization of Fermentation Conditions for Protease Production by Bacillus licheniformis with Enteromorpha prolifera as Carbon Source
WANG Ying
1, GAO Xiang
2, ZHOU Jun
1, ZHANG Chundan
1, LI Ye
1, WANG Zuzhong
1, YUAN Bei
1, DAI Juan
1, QIAN Qinlian
1, SU Xiurong
1,
*
(1. School of Marine Sciences, Ningbo University, Ningbo 315211, China; 2. Qiaosi Prison in Zhejiang, Hangzhou 310019, China)
Abstract:This work reports on the selection and optimization of culture conditions and medium components as well as their levels for improved production of protease by Bacillus licheniformis with Enteromorpha prolifera as a carbon source by the combined use of single factor method, Plackett-Burman design and response surface methodology (RSM). Initial medium pH, Enteromorpha prolifera concentration and NaH 2PO 4concentration were identified as the most significant factors that influence protease production. The levels of the three factors were optimized using RSM to be 6.66, 4.65% and 0.027 4%, respectively. Under the optimized conditions, the predicted values of protease activity was 66 354.70 U/g protein and an average value of 66 966.37 U/g protein was obtained from three replicate experiments, which was 3.68 times higher than that before optimization.
Key words:Bacillus licheniformis; Enteromorpha prolifera; fermentation conditions; protease; Plackett-Burman design
DOI:10.7506/spkx1002-6630-201607022
中图分类号:Q939.9
文献标志码:A
文章编号:1002-6630(2016)07-0117-06
引文格式:
王颖, 高翔, 周君, 等. 以浒苔为碳源的地衣芽孢杆菌发酵产蛋白酶条件优化[J]. 食品科学, 2016, 37(7): 117-122.
DOI:10.7506/spkx1002-6630-201607022. http://www.spkx.net.cn
WANG Ying, GAO Xiang, ZHOU Jun, et al. Optimization of fermentation conditions for protease production by Bacillus licheniformis with Enteromorpha prolifera as carbon source[J]. Food Science, 2016, 37(7): 117-122. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201607022. http://www.spkx.net.cn
收稿日期:2015-10-01
基金项目:国家科技星火计划项目(2010GA701063);浙江省重点社会发展项目(2009C03017-1)
作者简介:王颖(1990—),女,硕士研究生,研究方向为食品科学。E-mail:plyinger@126.com
*通信作者:苏秀榕(1956—),女,教授,博士,研究方向为食品科学与工程/生化与分子生物学。E-mail:suxiurong@nbu.edu.cn