刘春娟 1,边 鑫 2,赵士举 3
(1.吉林省经济管理干部学院现代农业学院,吉林 长春 130012;2.东北农业大学食品学院,黑龙江 哈尔滨 150030;3.河南农业大学理学院,河南 郑州 450002)
摘 要:以青霉菌作为酸乳腐败菌筛选具有抑制该菌能力的罗伊氏乳杆菌,并通过排除实验确定优势菌株的主要抑菌物质,再将优势菌株作为辅助发酵剂用于酸乳发酵,并对酸乳进行理化性质和感官品质的评定以及优势菌株在酸乳中生物防腐作用的探讨。结果表明,罗伊氏乳杆菌CICC6121是具有抑制青霉菌能力的优势菌株,其无细胞上清液中抑制青霉菌的主要活性成分为罗伊氏乳杆菌素;罗伊氏乳杆菌CICC6121在酸奶发酵过程中不会对保加利亚乳杆菌和嗜热链球菌产生拮抗作用,且所生产的酸乳与未接种CICC6121时的酸乳在理化性质和感官品质方面几乎无差别,在21 d的低温贮藏过程中,能够抑制酸乳中青霉菌的生长,减少因污染而造成酸乳的腐败变质。
关键词:酸乳腐败菌;抑菌作用;罗伊氏乳杆菌;酸乳感官品质;生物防腐
酸乳是一类营养价值高、具有保健作用的发酵乳制品,在改善乳糖不耐症、促进消化和调节肠道菌群等方面发挥着重要的作用。通常,酸乳的保质期大概在15 d左右,加入防腐剂的酸乳可延长至约21 d左右。然而,在实际情况下,酸乳的保质期通常会大打折扣,甚至在贮藏初期就会出现鼓盖、发霉以及大量乳清析出等现象。这种现象主要是由酸乳的后酸化作用和霉菌、酵母菌的污染两个原因造成的。酸乳在发酵完成后冷藏期间,某些乳酸菌会在低温条件下继续发酵产酸,导致酸乳凝胶结构松散,持水能力下降,使酸乳的品质变差;同时,酸乳在生产过程中会由于操作不当等原因,造成腐败菌对酸乳的污染,使酸乳在冷藏过程中发霉变质。这些腐败菌通常包括:青霉属、汉氏德巴利氏酵母属(Debaryomyces hansenii)、马克斯克鲁维酵母属(Kluyveromyces marxianus)、红酵母属(Rhodotorula mucilaginosa)、解脂耶氏酵母属(Yarrowia lipolytica)以及接合酵母属(Zygosaccharomyces bailii)等 [1]。其中,青霉菌是酸奶发霉变质的主要元凶。Jordano等 [2]分别调查了部分市场出售的酸奶样品,结果表明,青霉菌的污染比例达到70%以上。这些腐败微生物不仅破坏了酸乳的各种营养成分,而且还会在酸乳中生长繁殖,并产生霉菌毒素,对酸乳的品质和安全性产生影响。
目前,大多乳酸菌都是益生菌,它们分泌的一些代谢产物可以抑制某些细菌和真菌的生长,在生物防腐方面具有很好的应用潜力 [3],例如干酪乳杆菌 [4](抑制腐败菌)、植物乳杆菌 [5](抑制腐败菌)嗜酸乳杆菌 [6]、发酵乳杆菌 [7]、哈尔滨乳杆菌 [8]以及保加利亚乳杆菌 [9](抗后酸化)等在生物防腐方面的作用均已有相关报道。此外,某些乳酸菌的产酸能力相对较弱,有效地减弱后酸化作用。罗伊氏乳杆菌(Lactobacillus reuteri)为革兰氏阳性菌,天然存在于哺乳动物及鸟类的肠道中 [10]。目前,已有部分研究证实其可产生广谱抑菌物质,在生物防腐方面具有一定潜力 [11-12]。然而由于罗伊氏乳杆菌的来源以及功能不同,无法保证所有菌株都具有生物防腐能力。因此,本实验以青霉菌作为酸乳的腐败指示菌,筛选出抑菌能力较强的罗伊氏乳杆菌,并将其作为辅助发酵剂应用于酸乳的生产,对酸乳的理化性质和感官品质以及罗伊氏乳杆菌在酸乳中抑制腐败菌的能力进行研究,为微生物法延长酸乳保质期提供优势菌株。
1.1 菌株、试剂与培养基
罗伊氏乳杆菌(Lactobacillus reuteri)CICC6119、CICC6120、CICC6121、CICC6124、CICC6126、CICC6131、CICC6132、CICC6226 中国工业微生物菌种保藏管理中心;作为酸乳腐败指示菌的青霉菌(Penicillium)CFCC88688 中国林业微生物菌种保藏管理中心。
蛋白胨、胰蛋白胨、牛肉膏、酵母浸粉 北京奥博星生物技术有限责任公司;其他试剂均为分析纯。
罗伊氏乳杆菌采用改良MRS培养基进行培养,改良MRS培养基参照苏本宪 [13]的方法。青霉菌采用马铃薯葡萄糖琼脂培养基(potato dextrose agar,PDA)培养基进行培养,PDA培养基的配制参照边鑫等 [14]的方法。
1.2 仪器与设备
SW-CJ无菌操作台 苏州市吴江诚信净化设备厂;高速冷冻离心机 上海市离心机械研究所;XT5107-IB150生化培养箱 杭州雪中炭恒温技术有限公司;TQHZ-2002A全温度振荡培养箱 太仓市华美生化仪器厂;移液器 大龙医疗设备(上海)有限公司;高精度笔式酸度计 杭州联测自动化技术有限公司;乌氏黏度计 上海隆拓仪器设备有限公司。
1.3 方法
1.3.1 实验菌株的活化和培养
从8 株罗伊氏乳杆菌冻存管取适量菌液接种于改良MRS液体培养基中37 ℃下培养活化传代两次,每次培养24 h。将活化后的菌液于4 ℃冰箱保存备用。
从冻存管中取适量的青霉菌菌液接种于PDA液体培养基25 ℃培养5 d后,接种于PDA斜面培养基25 ℃培养5 d后于4 ℃冰箱保存备用。
1.3.2 具有抑制腐败菌活性的罗伊氏乳杆菌菌株的筛选
1.3.2.1 无细胞上清液的制备
8 株罗伊氏乳杆菌的无细胞上清液按照以下步骤进行制备:罗伊氏乳杆菌在改良MRS培养中37 ℃下培养24 h,取菌液于10 000 r/min、4 ℃下离心10 min,弃沉淀,得到上清液于0.22 μm滤膜过滤,即得罗伊氏乳杆菌无细胞上清液。
1.3.2.2 高抑制腐败菌能力菌株的筛选
采用牛津杯法 [15]进行抑菌活性的评定,具体方法为:向培养皿中倒入10 mL 1%琼脂培养基,待凝固后,再倒入10 mL含有1%酸乳腐败指示菌的PDA半固体培养基和麦芽汁半固体培养基,凝固后将2 个牛津杯置于培养基表面,轻轻压实使接触面无空隙。向其中1 个牛津杯中加入100 μL罗伊氏乳杆菌无细胞上清液,另1 个牛津杯中加入100 μL无菌水作为空白对照,分别在适宜的温度下进行培养。用游标卡尺对抑菌圈直径进行测量,其中实验结果需减去牛津杯直径。根据抑菌圈直径的大小筛选抑菌能力较强的罗伊氏乳杆菌菌株。
1.3.3 高抑制腐败菌能力菌株无细胞上清液中抑菌成分的确定
取高抑制腐败菌能力菌株无细胞上清液分成5 组,第1组用2 mol/L的NaOH调节pH值为7.0;第2组加入5 mg/mL的过氧化氢酶于37 ℃水浴2 h;第3组加入1 mg/mL蛋白酶K于37 ℃水浴2 h;第4组加入1 mg/mL木瓜蛋白酶于37 ℃水浴2 h,第5组不做任何处理作为空白对照。以上所有上清液经0.22 μmL滤膜过滤后,加入到牛津杯中进行抑菌实验,通过计算各组高抑制腐败菌能力菌株无细胞上清液的残留抑菌活性来确定主要的抑菌成分,公式如下。
1.3.4 高抑制腐败菌能力菌株在酸乳中的应用
1.3.4.1 含罗伊氏乳杆菌酸乳(简称R菌酸乳)的制备
取适量牛乳于三角瓶中,将酸奶发酵剂(保加利亚乳杆菌:10 8CFU/mL;嗜热链球菌:10 8CFU/mL)以3%的接种量接种于经过灭菌的牛乳中,同时取罗伊氏乳杆菌酸乳作为辅助发酵剂分别以1%、2%、3%、4%和5%的接种量接种于经过灭菌的牛乳中,42 ℃条件下发酵,当酸乳表面出现少量乳清后停止发酵,放入4 ℃冰箱冷藏24 h。
1.3.4.2 酸乳理化性质的测定和感官评价
酸乳在冷藏过程中pH值的变化采用高精度笔式酸度计进行测定。将酸度计经巴氏杀菌后打开开关,然后插入刚刚完成发酵的酸乳中,第1天先测定一次pH值,此后每7 d计数一次。酸乳的酸度滴定参照范瑞等 [16]方法进行测定。酸乳的表观黏度采用乌氏黏度计进行测定,具体操作参照Ozturk等 [17]的方法。酸乳的持水能力参照Lucey等 [18]方法进行测定,持水能力的计算公式如下。
式中:m 1为析出乳清的质量/g,m 2为发酵前原料乳和加入发酵剂的总质量/g。
酸乳的感官评价参照GB 19302ü2010《发酵乳》 [19],随机选择20 名评价员参与感官评价,分别从色泽(满分10 分)、滋味和气味(满分40 分)和组织状态(满分50 分)三方面进行评定。将所有评价员的评定结果去平均数即为酸乳的感官评分。
1.3.4.3 酸乳中乳酸菌活菌数的测定
酸乳中各乳酸菌活菌数的测定参照Marafon [20]和Saccaro [21]等的方法。首先取适量酸乳样品用无菌去离子水稀释适当的浓度,然后取100 μL稀释样品分别均匀涂布于改良MRS琼脂培养基和M17琼脂培养基上,37 ℃下培养48 h。根据菌体形态不同可区分改良MRS琼脂培养基上的保加利亚乳杆菌(菌落为淡白色)和罗伊氏乳杆菌(其菌落为灰色)并对其进行菌落计数,同时对M17琼脂培养基上的嗜热链球菌进行菌落计数。所得菌落数乘以稀释倍数即为乳酸菌的活菌数。
1.3.4.4 高抑制腐败菌能力菌株在酸乳中的抑菌实验
将含不同浓度罗伊氏乳杆菌CICC6121的酸乳无菌操作倒入已灭菌的培养皿中,然后放入4 ℃冰箱冷藏21 d,分别在第1、7、14、21天观察酸乳表面腐败菌生产情况,并进行菌落计数。另取含不同浓度CICC6121的酸乳,向其中接入1%的青霉菌(10 5CFU/mL左右)后混匀,将处理过的酸乳放入4 ℃冰箱冷藏21 d,分别在第1、7、14、21天观察酸乳表面腐败菌生产情况,并进行菌落计数。
1.4 数据处理
以上所有实验均重复3 次,结果用
 表示,图表用Excel 2010软件进行绘制;数据处理采用SPSS软件进行统计学分析。
表示,图表用Excel 2010软件进行绘制;数据处理采用SPSS软件进行统计学分析。
2.1 高抑制腐败菌能力菌株的筛选结果
酸乳腐败变质主要是由腐败菌污染而导致的,其中以青霉菌污染最为常见,因此,本研究选取青霉菌作为酸乳腐败指示菌,通过8 株罗伊氏乳杆菌对其的抑菌作用筛选出优势菌株,筛选结果如表1所示。
表1 8 株罗伊氏乳杆菌的无细胞上清液对青霉菌的抑菌效果
Table 1 Inhibitory effect of cell free supernatants of 8L. reuteri against Penicillium

酸乳腐败指示菌抑菌圈直径/mm CICC6119 CICC6120 CICC6121 CICC6124 CICC6126 CICC6131 CICC6132 CICC6226青霉菌CFCC 88688 5.05f0.12 7.36f0.10 11.84f0.07 3.25f0.23 4.11f0.08 4.12f0.14 5.63f0.25 6.53f0.22
由表1可知,8 株罗伊氏乳杆菌对青霉菌均有抑制作用,其中CICC6121的抑菌圈直径明显大于剩余7 株罗伊氏乳杆菌,达到(11.84f0.07) mm。因此,可以确定在提供的罗伊氏乳杆菌中,CICC6121为高抑制腐败菌能力菌株。
2.2 罗伊氏乳杆菌CICC6121的主要抑菌成分
为了确定罗伊氏乳杆菌CICC6121的主要抑菌成分,对其无细胞上清液分别调节不同pH值、加入5 mg/mL的过氧化氢酶、加入1 mg/mL蛋白酶K和加入1 mg/mL木瓜蛋白酶处理,残留的抑菌活性见表2。
表2 罗伊氏乳杆菌CICC6121主要抑菌物质的分析
Table 2 Analysis of antimicrobial substances from L. reuteri CICC6121

1 mg/mL木瓜蛋白酶组青霉菌CFCC 8868880.21f0.35 95.31f0.13 99.42f0.27 99.34f0.45酸乳腐败指示菌残留的抑菌活性/% pH 7组 5 mg/mL过氧化氢酶组1 mg/mL蛋白酶K组
以空白对照组残留的抑菌活性为100%,由表2可知,CICC6121无细胞上清液经中和处理后,抑菌活性有所降低,说明其产生的有机酸对青霉菌有一定的抑制作用;经过5 mg/mL的过氧化氢酶处理后的上清液抑菌活性变化不大,说明过氧化氢不是抑制青霉菌的有效成分;而无细胞上清液经1 mg/mL蛋白酶K和1 mg/mL木瓜蛋白酶处理后,抑菌活性几乎无任何变化,说明蛋白类物质不是主要的抑菌成分。此前有大量研究表明,罗伊氏乳杆菌素具有广谱抑菌活性 [22-23]。因此可以推断CICC6121抑制青霉菌的主要活性成分为罗伊氏乳杆菌素。
2.3 R菌酸乳的理化性质及感官品质
2.3.1 R菌酸乳的发酵时间、滴定酸度和pH值
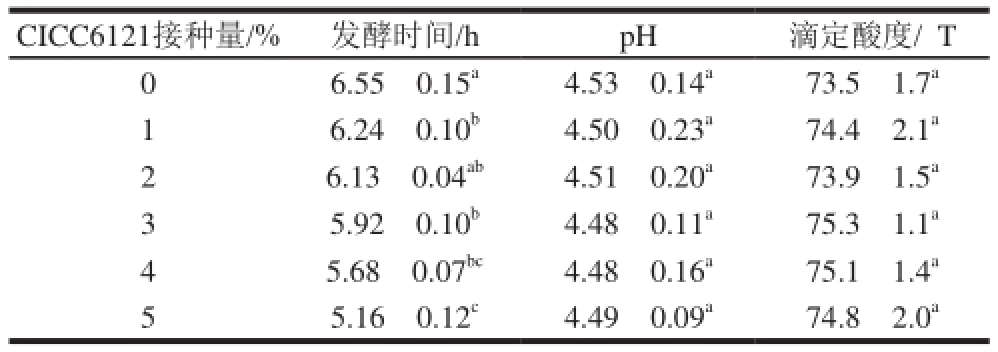
由于发酵剂接种量的不同,酸乳的发酵时间及相关参数可能也不尽相同。不同CICC6121接种量的酸乳所需的发酵时间以及发酵完成时的pH值和滴定酸度见表3。
表3 罗伊氏乳杆菌CICC6121不同接种量发酵酸乳的发酵时间、pH值、滴定酸度
Table 3 Fermentation time, pH, titratable acidity of yogurt at different inoculum sizes of L. reuteri CICC6121
注:同列小写字母不同表示差异显著(P<0.05)。下同。
CICC6121接种量/% 发酵时间/h pH 滴定酸度/eT 0 6.55f0.15 a4.53f0.14 a73.5f1.7 a1 6.24f0.10 b4.50f0.23 a74.4f2.1 a2 6.13f0.04 ab4.51f0.20 a73.9f1.5 a3 5.92f0.10 b4.48f0.11 a75.3f1.1 a4 5.68f0.07 bc4.48f0.16 a75.1f1.4 a5 5.16f0.12 c4.49f0.09 a74.8f2.0 a
由表3可知,随着CICC6121接种量的增加,酸乳的发酵时间逐渐缩短,其中接种5% CICC6121酸乳的发酵时间比接种1% CICC6121酸乳缩短了1 h左右,比未接种CICC6121酸乳的发酵时间缩短了将近1.5 h。此外,不同接种量发酵的酸乳的pH值和滴定酸度则没有显著性差异,且滴定酸度均在70 eT以上,符合酸乳国家标准GB 19302ü2010《发酵乳》 [19],以上结果表明,接种CICC6121缩短了酸乳的发酵时间,但未对酸乳的相关参数产生影响。
2.3.2 R菌酸乳在冷藏过程中pH值变化

图1 罗伊氏乳杆菌CICC6121不同接种量发酵的酸乳在4 ℃条件下冷藏21 d的pH值变化
Fig.1 Change in pH of yogurt fermented with different inoculum amounts of L. reuteri CICC6121 during 21 days storage at 4 ℃
由图1可知,酸乳随着冷藏时间的延长,pH值逐渐降低,说明酸乳在冷藏过程中产生了后酸化作用,导致体系的pH值的下降。此外,接种罗伊氏乳杆菌CICC6121所制酸乳的pH值显著高于空白组,说明罗伊氏乳杆菌CICC6121对酸乳在冷藏过程中的后酸化有一定的抑制作用。
2.3.3 R菌酸乳在冷藏过程中滴定酸度的变化
由表4可知,随着冷藏时间的延长,5 组酸乳的滴定酸度均有所升高,这是由于酸乳在冷藏过程中,乳酸菌继续利用乳糖产酸,导致体系的乳酸浓度增加,从而使滴定酸度逐渐上升。同时,研究发现,5%接种量酸乳的酸度的滴定酸度略低于其他接种量酸乳的滴定酸度,但差异不显著,说明罗伊氏乳杆菌CICC6121对酸乳在冷藏过程中的后酸化有一定的抑制作用。
表4 罗伊氏乳杆菌CICC6121不同接种量发酵的酸乳在4 ℃条件下冷藏21 d的滴定酸度变化
Table 4 Titratable acidity of yogurt fermented with different inoculum amounts of L. reuteri CICC6121 during 21 days of storage at 4 ℃
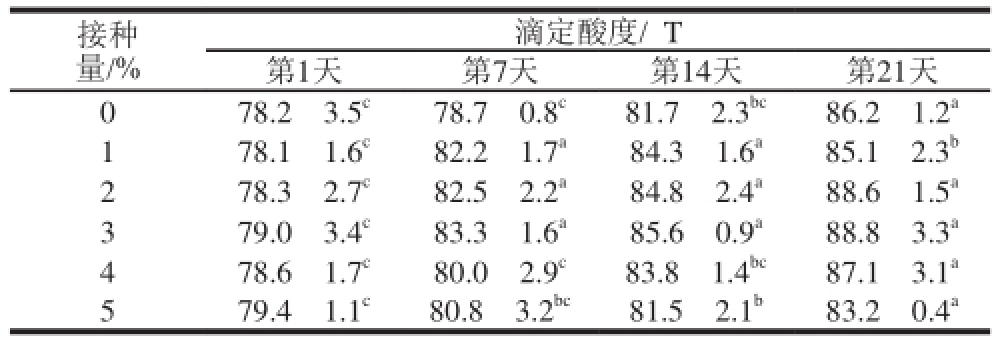
接种量/%滴定酸度/eT第1天 第7天 第14天 第21天0 78.2f3.5 c78.7f0.8 c81.7f2.3 bc86.2f1.2 a1 78.1f1.6 c82.2f1.7 a84.3f1.6 a85.1f2.3 b2 78.3f2.7 c82.5f2.2 a84.8f2.4 a88.6f1.5 a3 79.0f3.4 c83.3f1.6 a85.6f0.9 a88.8f3.3 a4 78.6f1.7 c80.0f2.9 c83.8f1.4 bc87.1f3.1 a5 79.4f1.1 c80.8f3.2 bc81.5f2.1 b83.2f0.4 a
2.3.4 R菌酸乳在冷藏过程中表观黏度的变化
表5 罗伊氏乳杆菌CICC6121不同接种量发酵的酸乳在4 ℃条件下冷藏21 d的表观黏度变化
Table 5 Apparent viscosity of yogurt fermented with different inoculum amounts of L. reuteri CICC6121 during 21 days of storage at 4 ℃
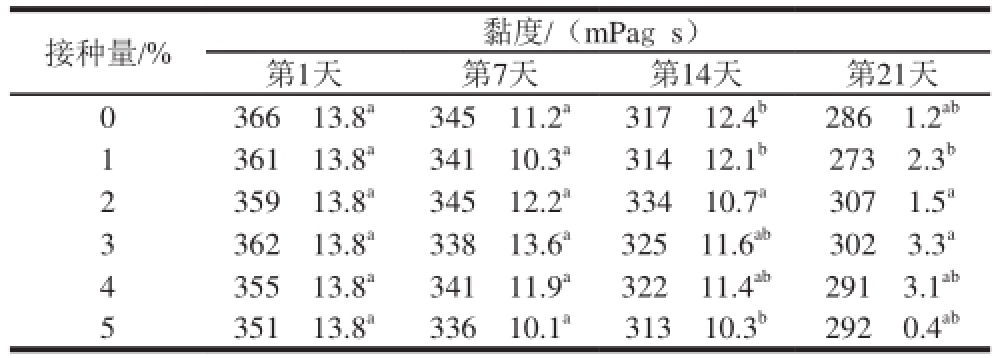
接种量/% 黏度/(mPags)第1天 第7天 第14天 第21天0 366f13.8 a345f11.2 a317f12.4 b286f1.2 ab1 361f13.8 a341f10.3 a314f12.1 b273f2.3 b2 359f13.8 a345f12.2 a334f10.7 a307f1.5 a3 362f13.8 a338f13.6 a325f11.6 ab302f3.3 a4 355f13.8 a341f11.9 a322f11.4 ab291f3.1 ab5 351f13.8 a336f10.1 a313f10.3 b292f0.4 ab
由表5可知,随着冷藏时间的延长,5 组酸乳的表观黏度均有所下降,这可能由于酸乳中的乳酸菌所产蛋白酶在发酵完成后继续发挥作用;同时随着后酸化程度的增加,使体系的pH值持续降低,这两方面使蛋白胶粒之间的亲和连接作用减弱,从而导致酸乳凝胶网络结构变得松散,体系黏度下降 [24-25]。另外,本课题组对罗伊氏乳杆菌CICC6121不同接种量发酵的酸乳的表观黏度进行了比较,结果表明,不同接种量的酸乳在同一时期的表观黏度差异不显著,说明罗伊氏CICC6121的接种量对酸乳的表观黏度影响不大。因此,可以根据需要选取适当的接种量进行酸乳的发酵。
2.3.5 R菌酸乳在冷藏过程中持水能力的变化
由表6可知,随着冷藏时间的延长,5 组酸乳的持水能力均有所下降,这可能由于酸乳的后酸化导致了体系的pH值持续降低,使酸乳凝胶网络结构变得松散,持水能力降低。同时对罗伊氏乳杆菌CICC6121不同接种量发酵的酸乳的持水能力进行了比较。结果表明,不同接种量的酸乳在同一时期的持水能力差异不显著,说明罗伊氏CICC6121的接种量对酸乳的持水能力影响不大。因此,可以根据需要选取适当的接种量进行酸乳的发酵。
表6 罗伊氏乳杆菌CICC6121不同接种量发酵的酸乳在4 ℃条件下冷藏21 d的持水能力变化
Table 6 Water-holding capacity of yogurt fermented with different inoculum amounts of L. reuteri CICC6121 during 21 days of storage at 4 ℃
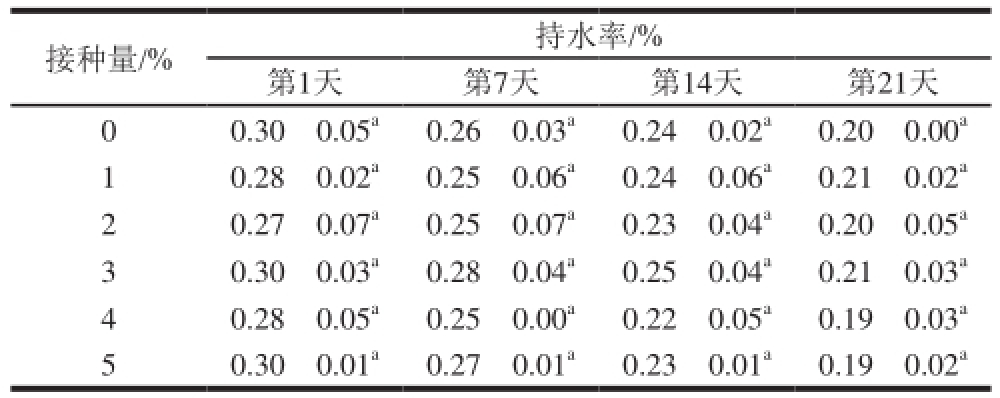
接种量/% 持水率/%第1天 第7天 第14天 第21天0 0.30f0.05 a0.26f0.03 a0.24f0.02 a0.20f0.00 a1 0.28f0.02 a0.25f0.06 a0.24f0.06 a0.21f0.02 a2 0.27f0.07 a0.25f0.07 a0.23f0.04 a0.20f0.05 a3 0.30f0.03 a0.28f0.04 a0.25f0.04 a0.21f0.03 a4 0.28f0.05 a0.25f0.00 a0.22f0.05 a0.19f0.03 a5 0.30f0.01 a0.27f0.01 a0.23f0.01 a0.19f0.02 a
2.3.6 R菌酸乳的感官评分
为了保证酸乳的口感、气味、味道等方面达到一定的品质,随机选取了20 名评价员对R菌酸乳进行了感官评定,根据1.3.4.2节中所述的标准对不同接种量酸乳打分,评价结果见图2。

图2 罗伊氏乳杆菌CICC6121不同接种量发酵酸乳的感官评分
Fig.2 Sensory evaluation of yogurt with different inoculum amounts of L. reuteri CICC6121
由图2可知,加入罗伊氏乳杆菌CICC6121的酸乳与空白组的感官评分基本相近,均在80~90 分之间,且不同接种量的酸乳之间感官评分的差异不显著,说明加入罗伊氏CICC6121后未对酸乳的感官品质产生影响,可作为发酵剂或辅助发酵剂用于酸乳的生产。
2.3.7 R菌酸乳冷藏过程中乳酸菌的生长情况
表7 接种1%R菌的酸乳在4 ℃条件下冷藏21 d过程中乳酸菌生长情况
Table 7 Growth of spoilage organisms in yogurt fermented with 1% L. reuteri CICC6121 during 21 days of storage at 4 ℃
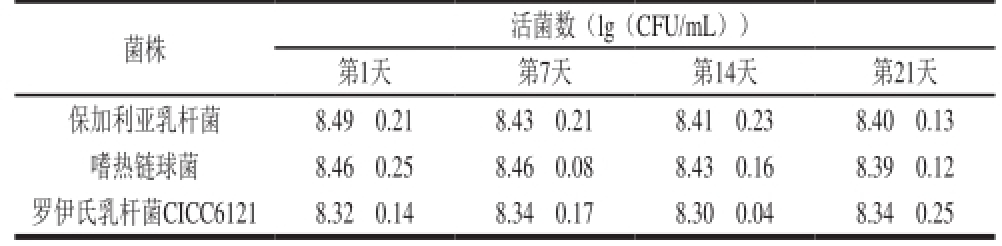
菌株 活菌数(lg(CFU/mL))第1天 第7天 第14天 第21天保加利亚乳杆菌 8.49f0.21 8.43f0.21 8.41f0.23 8.40f0.13嗜热链球菌 8.46f0.25 8.46f0.08 8.43f0.16 8.39f0.12罗伊氏乳杆菌CICC6121 8.32f0.14 8.34f0.17 8.30f0.04 8.34f0.25
由表7可知,接种1%R菌的酸乳在4℃下冷藏21d的过程中,保加利亚乳杆菌和嗜热乳杆菌的活菌数略有下降,而罗伊氏乳杆菌CICC6121的活菌数变化不大。由于酸奶在贮藏过程中,乳酸菌所产有机酸持续积累,导致pH值不断降低,保加利亚乳杆菌和嗜热乳杆菌的耐酸能力相对较差,活菌数量应大幅度下降。而在1%R菌酸奶中,二者的活菌数没有显著性变化,这可能与罗伊氏乳杆菌的后酸化能力较弱,延缓了体系的pH值的下降速率和程度有关。
2.3.8 R菌酸乳在冷藏过程中腐败菌的生长情况
将R菌酸乳置于平板中,盖盖后于4 ℃条件下冷藏21 d,在冷藏过程中腐败菌污染情况见表8。
表8 罗伊氏乳杆菌CICC6121不同接种量发酵的酸乳在4 ℃条件下冷藏21 d过程中腐败菌生长情况
Table 8 Growth of spoilage organisms of L. reuteri CICC6121-added yogurt with different inoculum amounts during 21 days storage at 4 ℃

接种量/% 腐败菌菌落数/个第1天 第7天 第14天 第21天0 0 0 2 3 1 0 0 0 0 2 0 0 0 0 3 0 0 0 0 4 0 0 0 0 5 0 0 0 0
由表8可知,加入罗伊氏乳杆菌CICC6121的酸乳在冷藏21 d后均没有腐败菌菌落的出现,而空白对照组则第14天出现2 个腐败菌菌落,当冷藏21 d时菌落数达到了3 个。结果表明,罗伊氏乳杆菌在酸乳中可有效抑制腐败菌的生长,说明在酸乳中添加适量罗伊氏乳杆菌CICC6121能在一定程度上延长酸乳的保质期。
表9 罗伊氏乳杆菌CICC6121不同接种量发酵的酸乳被青霉菌污染后在4 ℃条件下冷藏21 d的青霉菌生长情况
Table 9 Growth of Penicillium in artificially contaminated yogurt fermented with of different inoculum amounts of L. reuteri CICC6121 during 21 days storage at 4 ℃

接种量/% 青霉菌菌落数/个第1天 第7天 第14天 第21天0 0 2 3>3 1 0 0 0 0 2 0 0 0 0 3 0 0 0 0 4 0 0 0 0 5 0 0 0 0
由表9可知,空白对照组中,4 ℃条件下冷藏的R菌酸乳在第7天出现青霉菌菌落,而接种罗伊氏乳杆菌CICC6121的酸乳在4 ℃条件下冷藏21 d内均未出现青霉菌菌落。结果表明,罗伊氏乳杆菌CICC6121能抑制酸乳中的青霉菌,说明罗伊氏乳杆菌CICC6121的添加可有效延长酸乳的保质期。
本研究以青霉菌作为酸乳腐败菌筛选出具有抑制该菌的罗伊氏乳杆菌CICC6121,并确定该菌的主要抑菌物质为罗伊氏乳杆菌素;同时CICC6121对保加利亚乳杆菌和嗜热链球菌产生拮抗作用,可作为辅助发酵剂用于酸乳的生产。此外,接种CICC6121的酸乳与未接种该菌的酸乳在理化性质和感官品质方面几乎相近,在21 d的低温保藏过程中,可抑制青霉菌的生长,从而有效延长酸乳的保质期。
参考文献:
[1] MAYORAL M B, MARTIN R, SANZ A, et al. Detection of Kluyveromyces marxianus and other spoilage yeasts in yoghurt using a PCR-culture technique[J]. International Journal of Food Microbiology, 2005, 105(1): 27-34. DOI:10.1016/j.ijfoodmicro.2005.06.006.
[2] JORDANO R, MEDINA LM, SALMERON J. Contaminating mycoflora in fermented milk[J]. Journal of Food Protection, 1991, 54(2): 131-132.
[3] DALIE D K D, DESCHAMPS A M, RICHARD-FORGET F. Lactic acid bacteria-potential for control of mould growth and mycotoxins: a review[J]. Food Control, 2010, 21(4): 370-380. DOI:10.1016/ j.foodcont.2009.07.011.
[4] LI H J, LIU L, ZHANG S W. Potential use of Lactobacillus casei AST18 as a bioprotective culture in yogurt[J]. Food Control, 2013, 34(34): 675-680. DOI:10.1016/j.foodcont.2013.06.023.
[5] 王应东, 陈冲, 王海宽. 抗真菌乳酸菌的筛选及其对馒头和酸奶的防腐实验[J]. 天津科技大学学报, 2015, 30(2): 11-15. DOI:10.13364/ j.issn.1672-6510.20140058.
[6] CORTES-ZAVALETA O, LOPEZ-MALO A, HERNANDEZMENDOZA A, et al. Antifungal activity of lactobacilli and its relationship with 3-phenyllactic acid production[J]. International Journal of Food Microbiology, 2014, 173(3): 30-35. DOI:10.1016/ j.ijfoodmicro.2013.12.016.
[7] GEREZ C L, TORRES M J, VALDEZ G F D, et al. Control of spoilage fungi by lactic acid bacteria[J]. Biological Control, 2013, 64(3): 231-237. DOI:10.1016/j.biocontrol.2012.10.009.
[8] DELAVENNEA E, CLIQUETA S, TRUNET C, et al. Characterization of the antifungal activity of Lactobacillus harbinensis K.V9.3.1Np and Lactobacillus rhamnosus K.C8.3.1I in yogurt[J]. Food Microbiology, 2015, 45(PtA): 10-17. DOI:10.1016/j.fm.2014.04.017.
[9] 曹凤波, 王晓明, 周晶, 等. 两株弱后酸化保加利亚乳杆菌发酵性能的研究[J]. 食品工业科技, 2014, 35(21): 131-145. DOI:10.13386/ j.issn1002-0306.2014.21.020.
[10] TALARICO T L, CASAS I A, CHUNG T C, et al. Production and isolation of reuterin, a growth inhibitor produced by Lactobacillus reuteri[J]. Antimicrobial Agents and Chemotherapy, 1988, 32(12): 1854-1858. DOI:10.1128/AAC.32.12.1854.
[11] JUAN L A, JAVIER F, PILAR G, et al. Antimicrobial activity of reuterin in combination with nisin against food-borne pathogens[J]. International Journal of Food Microbiology, 2004, 95(2): 225-229. DOI:10.1016/j.ijfoodmicro.2004.03.009.
[12] VALENTINE C, CHRISTOPHE L, SABINE V, et al. Inhibitory activity spectrum of reuterin produced by Lactobacillus reuteri against intestinal bacteria[J]. BMC Microbiology, 2007, 7(1): 1-9. DOI:10.1186/1471-2180-7-101.
[13] 苏本宪. 乳杆菌对Staphylococcus aureus和Streptococcus pyogenes抑菌作用的研究[D]. 哈尔滨: 东北农业大学, 2014: 15-16.
[14] 边鑫, 吴非. 高产γ-氨基丁酸霉菌菌株的筛选及诱变育种[J]. 食品科学, 2012, 33(21): 213-216.
[15] WANG X, CHI Z, YUE L, et al. Purification and characterization of killer toxin from a marine yeast Pichia anomala YF07b against the pathogenic peast in Crab[J]. Current Microbiology, 2007, 55(5): 396-401. DOI:10.1007/s00284-007-9010-y.
[16] 范瑞, 许静, 顾宗珠, 等. 酸乳货架期内理化特性的研究[J]. 中国乳品工业, 2008, 36(8): 26-29. DOI:10.3969/ j.issn.1001-2230.2008.08.006.
[17] OZTURK B A, ONER M D. Production and evaluation of yogurt with concentrated grape juice[J]. Journal of Food Science, 1999, 64(3): 530-532. DOI:10.1111/j.1365-2621.1999.tb15077.x.
[18] LUCEY J A, MUNRO P A, SINGH H. Whey separation in acid skim milk gels made with glucono-δ-lactone: effects of heat treatment and gelation temperature[J]. Journal of Texture Studies, 1998, 29(4): 413-426. DOI:10.1111/j.1745-4603.1998.tb00813.x.
[19] GB 19302ü2010 发酵乳[S].
[20] MARAFON A P, SUMI A, GRANATO D, et al. Effects of partially replacing skimmed milk powder with dairy ingredients on rheology, sensory profiling, and microstructure of probiotic stirred-type yogurt during cold storage[J]. Journal of Dairy Science, 2011, 94(11): 5330-5340. DOI:10.3168/jds.2011-4366.
[21] SACCARO D M, TAMIME A Y, PILLEGGI A, et al. The viability of three probiotic organisms grown with yoghurt starter cultures during storage for 21 days at 4℃[J]. International Journal of Dairy Technology, 2009, 62(3): 397-404. DOI:10.1111/j.1471-0307.2009.00497.x.
[22] KULEASAN H, CAKMAKCI M L. Effect of reuterin, produced by Lactobacillus reuteri on the surface of sausages to inhibit the growth of Listeria monocytogenes and Salmonella spp.[J]. Food/Nahrung, 2002, 46(6): 408-410. DOI:10.1002/1521-3803(20021101)46:6<408::AIDFOOD408>3.0.CO;2-T.
[23] DEE H D. The influence of temperature, salt and pH on the inhibitory effect of reuterin on Escherichia coli[J]. International Journal of Food Microbiology, 2002, 72(3): 225-231. DOI:10.1016/S0168-1605(01)00637-7.
[24] 沈辉. 酸乳发酵凝乳过程中的理化性质和生物活性[J]. 无锡轻工大学学报, 2000, 19(5): 443-445. DOI:10.3321/ j.issn:1673-1689.2000.05.006.
[25] 郭清泉. 酸奶制品发生后酸化主要发酵剂菌确定及性质研究[J].食品与发酵工业, 2001, 28(14): 24-27. DOI:10.3321/j.issn:0253-990X.2002.04.006.
Screening of Lactobacillus reuteri Inhibiting Spoilage Organisms and Its Application to Yoghourt Production
LIU Chunjuan
1, BIAN Xin
2, ZHAO Shiju
3
(1. College of Modern Agriculture, Jilin Province Economic Management Cadre College, Changchun 130012, China; 2. College of Food Science, Northeast Agricultural University, Harbin 150030, China; 3. College of Sciences, Henan Agricultural University, Zhengzhou 450002, China)
Abstract:Lactobacillus reuteri with antimicrobial activity against Penicillium as spoilage bacteria in yoghurt was screened and the major antimicrobial substances produced by the dominant strain were determined by exclusion tests. Then, the dominant strain was applied as an adjunct culture to fermented milk. Meanwhile, physical and chemical properties and sensory quality of the yoghurt as well as the antiseptic effect of the dominant strain on it were studied. Results showed that Lactobacillus reuteri CICC6121 was the dominant strain against Penicillium. The antimicrobial substance in cellfree supernatant was reuterin. CICC6121 did not show any antagonism against Lactobacillus bulgaricus or Streptococcus thermophilus during yoghurt fermentation. The physicochemical properties and sensory quality of yoghurt inoculated and not inoculated with CICC6121 were nearly the same. During 21 days of cold storage, CICC6121 could inhibit the growth of Penicillium significantly and avoid yoghurt spoilage via contamination.
Key words:yoghourt spoilage organisms; bacteriostatic effect; Lactobacillus reuteri; sensory quality of yoghourt; biopreservation
DOI:10.7506/spkx1002-6630-201607029
中图分类号:TS252.54
文献标志码:A
文章编号:1002-6630(2016)07-0157-06
引文格式:
刘春娟, 边鑫, 赵士举. 具有抑制腐败菌能力的罗伊氏乳杆菌筛选及在酸乳生产中的应用[J]. 食品科学, 2016, 37(7): 157-162. DOI:10.7506/spkx1002-6630-201607029. http://www.spkx.net.cn
LIU Chunjuan, BIAN Xin, ZHAO Shiju. Screening of Lactobacillus reuteri inhibiting spoilage organisms and its application to yoghourt production[J]. Food Science, 2016, 37(7): 157-162. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201607029. http://www.spkx.net.cn
收稿日期:2015-07-08
作者简介:刘春娟(1977—),女,副教授,博士,研究方向为乳品加工。E-mail:58412681@qq.com