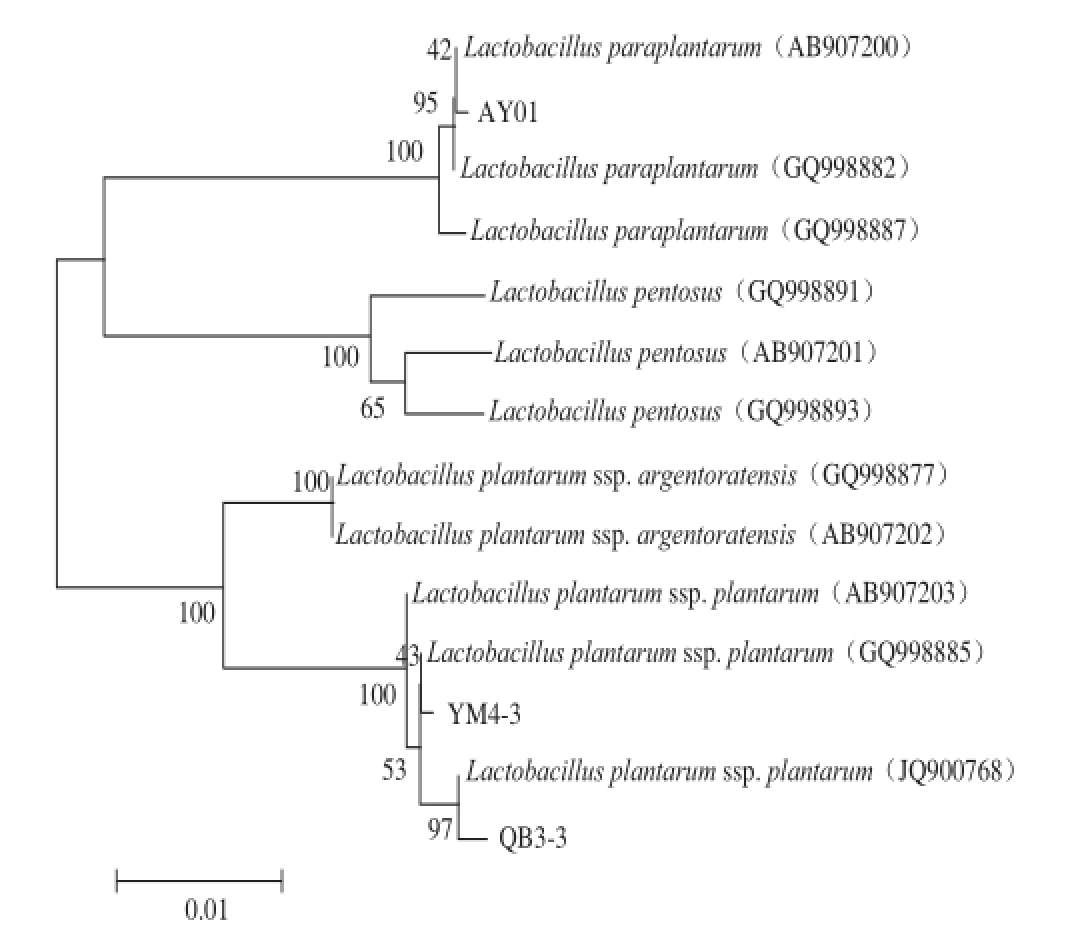
图
1 dnaKdnaK基因序列比对的植物乳杆菌亚种的系统进化树
Fig. 1 Phylogenetic tree for L. plantarum subspecies based on dnaK gene sequence alignment
李强坤,柳陈坚,张园莲,常林杰,罗义勇,李晓然*
(昆明理工大学生命科学与技术学院,云南 昆明 650500)
摘 要:通过牛津杯法筛选得到几株高抑菌活性的植物乳杆菌,使用蛋白酶K和NaOH对这几株杆菌的发酵液上清进行处理,以大肠杆菌O157:H7和金黄色葡萄球菌为指示菌,发现有机酸为主要抑菌物质,但是调节pH值至6.5后的发酵上清液可以对大肠杆菌有较好的抑菌效果。其中植物乳杆菌QB3-3具有长达96 h的持续抑菌效果。电子显微镜观察显示,植物乳杆菌抗病原菌的主要机理为破坏细胞结构,导致病原菌菌体形态改变,内容物流出,致使细胞死亡。
关键词:植物乳杆菌;有机酸;抑菌
食源性病原菌是指能在摄食过程中引起感染或中毒的一类微生物 [1],它广泛存在于食物、环境以及人和动物肠道内,能引起食物腐败、食源性腹泻等一系列卫生问题 [2-3]。使用抗生素与化学防腐剂可有效控制与杀灭食源性病原菌,然而,使用诸如苯甲酸(钠)、丙酸盐、对羟基苯甲酸酯类、硝酸盐、山梨酸(钾)等化学防腐剂不但会造成菌株的耐药性不断出现,同时也容易引起人的致畸与致癌等危害 [4-5],给人类健康造成严重威胁。随着我国国民经济的不断发展,人民生活水平的不断提高,食品安全受到人们越来越多的关注,食品污染造成的疾病已经成为全球性的公共卫生问题,而食源性病原菌污染是引起食品安全问题的主要原因 [6]。生物防腐剂对人体健康造成的危害较小,并且具有对食品营养成分及感官性状影响较小等优势 [7-9],同时还能减少防腐费用及有效延长产品保质期。此外,生物防腐剂还可避免化学防腐中出现的食物链的抗生素耐药性问题 [10]。因此,生物防腐不但受到研究者们的关注与重视,同时也深受普通百姓青睐。
乳酸菌是一类能将碳水化合物(主要指葡萄糖)发酵产生乳酸等有机酸的细菌。因其能产生有机酸、细菌素、环二肽、短链脂肪酸等多种抑菌活性物质 [11-12],故能有效抑制各类病原菌的生长繁殖,从而达到维持机体微生态平衡,发挥相应的益生功能 [13]。乳酸菌是被公认为安全(generally recognized as safe,GRAS)的食品级微生物(food-grade microorganisms)。植物乳杆菌(Lactobacillus plantarum)属于芽孢杆菌纲乳杆菌属(Lactobacillus) [14],通常存在于发酵蔬菜与果汁中,同时也发现其存在于传统发酵乳制品与肉类中,其发酵的食品被公认为功能性食品 [15]。植物乳杆菌能通过胃并定殖于肠道,发挥其调节肠道菌群平衡、免疫调节、降低血清胆固醇、降血压和抗氧化等益生作用 [15]。
地处西南低纬度地区的云南省因其多民族的文化背景,造就了独特的饮食风俗,从而富有多样的传统发酵食品资源,如乳饼、豆豉。前期研究表明,在云南传统发酵豆豉与乳饼中存在大量乳酸菌 [16],本实验从中筛选出3 株对食源性病原菌具有高效抑菌效果的植物乳杆菌,初步探究该乳酸菌的抗食源性致病菌的机理。
1.1 材料、试剂与培养基
从云南传统发酵食品(乳饼、豆豉)分离得到3株具有高效抑菌功能的植物乳杆菌AY01、YM4-3、QB3-3菌株、食源性大肠杆菌O157:H7、金黄色葡萄球菌,保存于昆明理工大学生命科学与技术学院应用微生物实验室。
琼脂 BIOSHARP公司;蛋白酶K 德国MERCK公司;其他试剂均为分析纯。
MRS培养基(g):蛋白胨10.0、Lab-lemco poweder 8.0、酵母提取物4.0、葡萄糖20.0、磷酸氢二钾2.0、醋酸钠5.0、柠檬酸三铵2.0、七水硫酸镁0.2、四水硫酸锰0.05、Tween-80 1 mL。加蒸馏水定容至1 L,121 ℃灭菌15 min,冷却后4 ℃保存备用。
脑心浸液肉汤(brian heart infusion,BHI)培养基(g):蛋白胨 10.0、牛肉浸粉3.0、氯化钠5.0、葡萄糖1.0,调pH值至7.4±0.1(25 ℃),加蒸馏水定容至1 L,121 ℃灭菌15 min,冷却后4 ℃保存备用。
肉汤(LB)培养基(g):蛋白胨 10.0、酵母粉 5.0、氯化钠 5.0、葡萄糖1.0,调pH值至7.1±0.1(25 ℃),加蒸馏水定容至1 L,121 ℃灭菌15 min,冷却后4 ℃保存备用。
1.2 仪器与设备
ZHWY-200D恒温培养振荡器 中国上海智城公司;ABI 7200 PCR仪 美国Applied Biosystems公司;UV-4802H紫外-可见分光光度计 尤尼柯(上海)仪器有限公司;电镜、FD-5-12真空冷冻干燥机 美国SIM公司;SX-500高压灭菌锅 日本TOMY公司;K3-18高速离心机 德国Sartorius公司;DYCP-31DN电泳仪美国BIO-RAD公司;SYIGLHR/1206凝胶成像系统英国Ingenius公司;BX-51研究级生物显微镜 日本Olympus公司;GHP-9160恒温培养箱 中国恒科学仪器有限公司。
1.3 方法
1.3.1 植物乳杆菌DNA的分离纯化
将冷冻保存的菌株解冻后按体积分数4‰接种量接种至MRS肉汤培养基中,35 ℃静置培养24 h,吸取1.5 mL菌液置于1.5 mL离心管中,4 ℃、8 000 r/min离心10 min,弃去上清。然后加入567 μL TE缓冲液、3 μL蛋白酶K(20 mg/mL)、30 μL 10%十二烷基硫酸钠(sodium dodecyl sulfate, sodium salt,SDS),充分混匀后,置于37 ℃金属浴孵育1 h。孵育结束后,加入80 μL十六烷基三甲基溴化铵(hexadecyl trimethyl ammonium bromide,CTAB)裂解液与100 μL 5 mol/L NaCl溶液,充分混匀后放入60 ℃金属浴孵育15 min。随后加入780 μL Tris-饱和酚充分振荡混匀,4 ℃、15 000 r/min离心10 min,取上清液,加入与上清液等体积的氯仿-异戊醇(24∶1,V/V),充分振荡混匀,4℃、15 000 r/min离心10 min后取上清液,加入0.6倍上清液体积的冷异丙醇,充分混匀后静置于-80 ℃条件下3 h。随后取出低温解冻,4 ℃、18 000 r/min离心15 min,弃去上清液,收集DNA沉淀,加入500 μL 70%的乙醇洗液洗脱2 次,4 ℃、18 000 r/min离心15 min后弃去上清液,置于室温使DNA沉淀充分干燥后,加入40 μL TE缓冲液。取5 μL DNA提取液与Loading Buffer混合进行1%琼脂糖凝胶电泳以检测基因组DNA,并选择检测到基因组DNA条带的样品,-80 ℃冷冻保存备用。
1.3.2 dnaK基因的PCR扩增反应及序列分析
采用细菌dnaK基因通用引物,用所提取的基因组DNA为模板进行扩增。dnaK通用引物核苷酸序列为:Lpdnak-500F3(5’-CCGTTCTTRTCRATRTCRAA-3’);Lpdnak-1710R5(5’-GAAAYYCAAGTYGGHGAAGT-3’)。用聚合酶链式反应(polymerase chain reaction,PCR)试剂盒进行扩增,PCR扩增反应程序为:94 ℃预变性5 min;94 ℃变性1 min,58 ℃退火1 min,72 ℃延伸1.5 min,35 个循环;72 ℃延伸7 min。PCR反应完成后,将扩增产物经1 g/mL琼脂糖凝胶电泳检测,电泳后将凝胶浸泡于含有20 μg/mL EB的TAE缓冲液中10 min,之后于紫外灯216 nm波长下照射,观察条带是否呈现橙色。将PCR产物纯化后,由上海生工生物工程股份有限公司进行核苷酸序列分析,提交至美国国家生物技术信息中心(National Center for Biotechnology Information,NCBI),用BLAST分析工具(http://www.ncbi.nlm.nih. gov/BLAST)将核苷酸序列与GenBank中已知核苷酸序列进行同源性对比。若被检菌株与参考菌株核苷酸序列同源性大于97%,将被认为是同一个菌种。最后用MEGA6.05软件的Neighbor-Joining法进行比对分析和建立系统发育树,Bootstrap重复1 000 次。
1.3.3 植物乳杆菌发酵上清液的制备
解冻保存的植物乳杆菌AY01、YM4-3、QB3-3,将浓度为1×10 9CFU/mL的植物乳杆菌菌液按体积分数4‰的接种量分别接种至灭菌的MRS肉汤培养基中,37 ℃恒温静止培养24 h后,再将其按体积分数4‰的接种量接种至300 mL锥形瓶中扩大培养72 h,随后4 ℃、12 000 r/min离心10 min后吸取上清液,分别将各菌株150 mL培养上清液分成3 等份,每种菌的3等份发酵上清液分别进行以下处理:1 份加入50 mg的蛋白酶K,终质量浓度为1 mg/mL,45 ℃水浴3 h;1 份加入NaOH溶液调pH值至6.5;1 份不做任何处理。将经上述处理的上清液用真空冷冻干燥法进行浓缩,浓缩后用0.9%生理盐水将粉末溶解为终质量浓度为0.5 mg/mL的溶液,最后用0.22 μm滤膜过滤除菌后,备用。
1.3.4 病原指示菌的制备
将-80 ℃冷冻保存的大肠杆菌O157:H7、金黄色葡萄球菌取出并迅速解冻,按体积分数4‰的接种量分别将其接种至5 mL LB液体培养基和BHI液体培养基,37 ℃,150 r/min振荡培养15 h,同时控制光密度值(optical density,OD)在0.2~0.8范围内。将培养好的菌液稀释后进行平板计数。大肠杆菌的菌落浓度为1.95×10 9CFU/mL,金黄色葡萄球菌的菌落浓度为2.83×10 9CFU/mL,再将活化好的病原菌菌液进行梯度稀释,用于抑菌实验。
采用牛津杯法进行抑菌实验,在底层平板中倒入10 mL 2%普通琼脂培养基,待其凝固后,按等边三角形放置3 个牛津杯。待上层培养基(大肠杆菌为LB琼脂培养基,金黄色葡萄球菌为BHI琼脂培养基)冷却至50 ℃左右时,取20 μL病原指示菌稀释液倒入25 mL 1%的上层琼脂培养基中,使大肠杆菌的终菌落总数为1.56×10 5CFU/mL,金黄色葡萄球菌的终菌落总数为2.26×10 5CFU/mL,摇匀后倒板。上层培养基凝固后取出牛津杯,在3 个孔中分别加入50、100、150 μL的抑菌物质(即植物乳杆菌发酵上清液),再用0.9%生理盐水定容至每孔150 μL,置于4 ℃直至抑菌液体完全扩散至琼脂培养基中,再置于37 ℃培养16 h,观察并测定抑菌圈的直径,每个实验做3 组平行。
病原指示菌敏感度的判定标准:抑菌圈直径>20 mm为极度敏感;抑菌圈直径15~20 mm为高度敏感;抑菌圈直径l0~15 mm为中度敏感;抑菌圈直径<10 mm为低度敏感;无抑菌圈为不敏感 [17]。
1.3.5 植物乳杆菌QB3-3菌株抑菌效果的实时监测
将50 μL活化的大肠杆菌O157:H7、金黄色葡萄球菌菌液添加至450 μL生理盐水中,吸取20 μL病原菌稀释液添加至5 mL相应的液体培养基中,分别向病原菌液体培养基中添加200、400、600 μL植物乳杆菌QB3-3发酵上清液,另外,将未添加任何抑菌溶液的病原菌菌液做为空白对照,在150 r/min,37 ℃的条件下振荡培养,前12h每隔1 h测定其在600nm波长处的OD值,之后每隔12 h测一次。
1.3.6 植物乳杆菌产生的抑菌物质对指示菌抑菌作用的电子显微镜观察
吸取200 μL植物乳杆菌QB3-3菌株发酵上清液至经过培养3 h且初始大肠杆菌浓度为1×10 9CFU/mL、5 mL的LB液体培养基中,于37 ℃、200 r/min振荡培养15 h;将200 μL 0.9%生理盐水添加至相同条件的LB液体培养基中作为阴性对照。培养结束后,4 ℃,3 000 r/min离心10 min取沉淀,用0.75%生理盐水洗脱菌体2 次,用1.5 mL固定液将菌体固定。用扫描电镜与透射电镜观察病原菌形态变化,探讨植物乳杆菌源抑菌物质的抑菌机理。
采用同样方法分析植物乳杆菌产生的抑菌物质对金黄色葡萄球菌的抑菌机理。
2.1 3 株植物乳杆菌亚种的鉴定
植物乳杆菌群包括Lb. paraplantarum、Lb. pentosus 2个种和Lb. plantarum ssp. plantarum、Lb. plantarum ssp. argentoratensis 2 个亚种,由于16S rRNA基因同源性较高,不能将植物乳杆菌鉴定到种或亚种的水平,而乳酸菌含有的编码适应酸环境的伴侣蛋白DnaK的dnaK基因常用于亚种的鉴定 [18]。通过对植物乳杆菌AY01、YM4-3、QB3-3的dnaK基因的核苷酸序列进行分析和同源性比对发现,从云南省石林县传统发酵乳饼的发酵酸水中分离得到的植物乳杆菌AY01菌株属于Lb. paraplantarum(序列同源性为99%),而分别从云南省丘北县与易门县传统发酵豆豉分离得到的菌株QB3-3与YM4-3和Lb. plantarum与Lb. plantarum ssp. plantarum的同源性均高于99%,比对结果见图1。由此可以将这2 株植物乳杆菌鉴定为植物乳杆菌植物乳杆亚种。
图
1 dnaKdnaK基因序列比对的植物乳杆菌亚种的系统进化树
Fig. 1 Phylogenetic tree for L. plantarum subspecies based on dnaK gene sequence alignment
2.2 植物乳杆菌发酵上清液的抑菌活性
3 株植物乳杆菌的抑菌实验表明,发酵上清原液与经蛋白酶K处理上清液对2 株病原菌均有抑菌作用(表1),对大肠杆菌的抑菌效果明显优于金黄色葡萄球菌。经NaOH调节pH值为6.5后,除添加高剂量(150 μL)植物乳杆菌YM4-3时,对金黄色葡萄球菌呈现微弱抑菌活性外,其余2 株植物乳杆菌对上述2 株食源性致病菌均未呈现抑菌效果。综合分析抑菌效果,植物乳杆菌QB3-3抑菌效果最好,其次是植物乳杆菌YM4-3。3 株植物乳杆菌的发酵上清液均显示较佳的抑菌效果,但对2 株病原菌产生抑菌效果所需的发酵上清液体积不同,100 μL的培养上清液即对大肠杆菌产生较好的抑菌效果,而150 μL的培养上清液才对金黄色葡萄球菌产生较好的抑菌效果。
表1 植物乳杆菌发酵上清液对食源性病原菌的抑菌效果
Table 1 Antimicrobial effect ofL. plantaarruumm against foodborne pathogennss

注:-. 无抑菌活性;抑菌圈直径:+. <10 mm,++. 抑菌圈直径l0~15 mm,+++. 15~20 mm,++++. >20 mm。
种类大肠杆菌 O157:H7金黄色葡萄球菌50 μL100 μL150 μL50 μL100 μL150 μL YM4-3(pH 6.5)-----+YM4-3(蛋白酶K)+++++++++++++++YM4-3原液 ++++++++++++++++AY01(pH 6.5)------AY01(蛋白酶K)+++++++++-+++AY01原液+++++++++-+++QB3-3(pH 6.5)------QB3-3(蛋白酶K)++++++++++++++QB3-3原液+++++++++++++++++
植物乳杆菌的抑菌物质主要分为蛋白类和非蛋白类物质,蛋白类抑菌物质主要为细菌素,是一种核糖体合成的抗菌肽,这些化合物可以作为安全的天然食品生物防腐剂 [19]。但细菌素并不是这3 株植物乳杆菌的主要抑菌物质 [20-21]。乳酸菌产生的非蛋白类物质主要为有机酸,大多数乳酸菌发酵碳水化合物生成乳酸、乙酸和苹果酸等有机酸,使环境pH值降低,抑制病原菌的生长,同时确保自身生长繁殖不受影响 [22]。植物乳杆菌的抑菌效果主要依赖其代谢产物,其中,有机酸就是其中的主要抑菌物质。
2.3 植物乳杆菌QB3-3抑菌效果的实时监测
根据3 株植物乳杆菌抑菌实验,选择抑菌效果最好的QB3-3进行长时间抑菌实时监测。监测数据通过软件Prism 6作图分析得图2、3。植物乳杆菌QB3-3在对大肠杆菌和金黄色葡萄球菌长达96 h的抑菌过程中,发酵原液的抑菌效果最佳,经蛋白酶K处理的上清液抑菌效果次之,调节pH值至6.5的上清液抑菌效果最差。

图2 植物乳杆菌 QB3-3对大肠杆菌O157:H7抑菌效果的实时监测
Fig. 2 Real-time monitoring of the antimicrobial effect of L. plantarum QB3-3 against E. coli O157:H7

图3 植物乳杆菌QB3-3对金黄色葡萄球菌抑菌效果的实时监测
Fig. 3 Real-time monitoring of the antimicrobial effect of L. plantarum QB3-3 against S. aureus
值得注意的是,当发酵上清液为600 μL时,即使pH值调至6.5,也对大肠杆菌有很好的抑菌效果。在Tulini等 [23]的研究中,植物乳杆菌所产生的细菌素对革兰氏阴性细菌不产生抑菌作用,而在本实验中,调节pH值后的发酵上清液主要的抑菌成分应为细菌素等蛋白质类抑菌物质,在较高添加量时表现出对大肠杆菌很好的抑制作用,说明研究细菌素的抑菌效果时,需对发酵上清液进行浓缩。并且,应对植物乳杆菌QB3-3所产的细菌素做更进一步的研究。
2.4 植物乳杆菌抑制致病菌的电子显微镜观察


图4 抑菌物质处理后的食源性病原菌电子显微镜图
Fig. 4 Photomicrographs of three L. plantarum strains with antibiotic susceptibility to E. coli O157:H7 and S. aureus
植物乳杆菌产生的抑菌物质对大肠杆菌的抑菌效果最好,并能持续较长时间,推测与革兰氏阴性菌和阳性菌的细胞壁不同有关。由图4可知,抑菌物质处理后的实验组和未经处理的对照组中,大肠杆菌菌体表面存在明显差异:1)正常生长状态的大肠杆菌菌体表面光滑、无空洞现象;经抑菌物质处理后的大肠杆菌菌体表面有黏丝状物质出现,菌体表面有褶皱并伴随有空洞。2)正常生长的金黄色葡萄球菌表面均匀光滑,无缺口;抑菌物质处理后的金黄色葡萄球菌细胞的内壁和外壁空间距离拉大,内壁浓缩并挤压细胞内容物,致使细胞壁出现裂口,内容物流出致死。在以前的研究中,Dieuleveux等 [24]用扫描电镜观察经苯乳酸作用前后的Listeria monocytogenes的超微结构后推测,苯乳酸的作用位点为细胞壁,与溶菌酶的抗菌机理相似,不同于Nisin等细菌素作用于细胞膜。袁景环等 [25]研究表明,苯乳酸具有杀菌作用,同时有一定的溶菌作用。通过扫描电镜和透射电镜观察显示,经苯乳酸作用后的致病菌的菌体形态发生明显改变,部分菌体破裂,细胞质流出。这些研究与本研究结果相似,因此,可以推测有机酸对细菌的作用靶点主要为细胞壁,而其具体的作用位点和抗菌机理有待进一步研究。
通过对dnaK基因序列比对分析,确定植物乳杆菌AY01为Lb. paraplantarum(序列同源性为99%),YM4-3和QB3-3均为Lb. plantarum ssp. plantarum。
实验分别使用植物乳杆菌AY01、YM4-3、QB3-3的发酵上清液,NaOH处理后的发酵上清液、蛋白酶K处理后的发酵上清液,进行对病原微生物大肠杆菌O157:H7和金黄色葡萄球菌的抑菌实验。结果表明,这3 株菌的主要抑菌物质为有机酸,其中菌株YM4-3和QB3-3具有微弱的非酸类抑菌物质。电子显微镜结果显示,经发酵液处理后的病原菌菌体结构发生变化,内容物流出,可见这3 株植物乳杆菌抗菌主要机理为破坏病原菌细胞结构,细胞内物质流出,致使细胞死亡。这些植物乳酸菌可作为优良的菌株,经改造后用于食品的防腐,具有良好的应用前景。
参考文献:
[1] LU X, YI L H, DANG J, et al. Purification of novel bacteriocin produced by Lactobacillus coryniformis MXJ 32 for inhibiting bacterial foodborne pathogens including antibiotic-resistant microorganisms[J]. Food Control, 2014, 46: 264-271. DOI:10.1016/ j.foodcont.2014.05.028.
[2] 孙香云, 刘学双, 高悦. 秦皇岛市北戴河区引起食源性腹泻的病原菌[J]. 职业与健康, 2013, 29(18): 2332-2334. DOI:10.13329/j.cnki. zyyjk.2013.18.033.
[3] 陶景渊. 浅谈急性肠胃炎症状及治疗方法[J]. 求医问药(学术版), 2012, 10(7): 267.
[4] 刘付香, 李玲, 梁炫强. 生物防治黄曲霉毒素污染研究进展[J]. 中国生物防治, 2010 (1): 96-101.
[5] 任广鸣. 化学防腐剂对豆腐贮存质量和营养成份的影响[J]. 食品科学, 1988, 9(12): 41-44.
[6] 王晓, 朱珠英, 张素燕, 等. 食品中食源性病原菌污染现况研究[J]. 浙江预防医学, 2011, 23(5): 12-16. DOI:10.3969/ j.issn.1007-0931.2011.05.004.
[7] 沈德中. 生物防治—防止果蔬腐烂的新途径[J]. 食品科学, 1990, 11(1): 55-58.
[8] 卢剑. 微生物防腐剂及其在食品工业中的应用[J]. 食品科学, 2005, 26(8): 453-457. DOI:10.3969/j.issn.1008-5467.2003.09.012.
[9] 樊星, 张昊, 郭慧媛, 等. 乳酸菌抑菌功能的研究进展[J]. 中国乳业, 2012(9): 52-54. DOI:10.3969/j.issn.1671-4393.2012.09.014.
[10] 张小平, 顾瑞霞. 益生菌发酵牛初乳对致病菌抑制作用的研究[J]. 中国乳品工业, 2005, 33(3): 10-12. DOI:10.3969/ j.issn.1001-2230.2005.03.003.
[11] SCHNÜRER J, MAGNUSSON J. Antifungal lactic acid bacteria as biopreservatives[J]. Trends in Food Science & Technology, 2005, 16(1/2/3): 70-78. DOI:10.1016/j.ti fs.2004.02.014.
[12] REIS J A, PAULA A T, CASAROTTI S N, et al. Lactic acid bacteria antimicrobial compounds: characteristics and applications[J]. Food Engineering Reviews, 2012, 4(2): 124-140. DOI:10.1007/s12393-012-9051-2.
[13] 杨玉红. 乳酸菌的抑菌活性及在肉品保鲜中的应用[J]. 肉类工业, 2013(6): 50-54. DOI:10.3969/j.issn.1008-5467.2013.06.016.
[14] 洪伟, 薛正莲. 1株植物乳杆菌的抑菌作用及其生物学特性的研究[J]. 生物学杂志, 2014(6): 51-54. DOI:10.3969/ j.issn.2095-1736.2014.06.051.
[15] 倪冬冬, 李洪军, 贺稚非. 新疆药桑果乳杆菌发酵饮料研制[J].食品工业科技, 2014, 35(24): 208-212. DOI:10.13386/ j.issn1002-0306.2014.24.036.
[16] 熊骏, 韩瑞娜, 张忠华, 等. 豆豉中高效抑菌活性乳酸菌的筛选及其抑菌研究[J]. 中国微生态学杂志, 2011, 23(6): 485-490.
[17] 张文, 韩广泉, 王凯, 等. 乳酸菌对中草药的发酵及抑菌活性检测[J].饲料博览, 2014(1): 4-7.
[18] HUANG C H, LEE F L. The dnaK gene as a molecular marker for the classifi cation and discrimination of the Lactobacillus casei group[J]. Antonie Van Leeuwenhoek, 2011, 99(2): 319-327. DOI:10.1007/ s10482-010-9493-6.
[19] GUINANE C M, COTTER P D, HILL C, et al. Microbial solutions to microbial problems; lactococcal bacteriocins for the control of undesirable biota in food[J]. Journal of Applied Microbiology, 2005, 98(6): 1316-1325. DOI:10.1111/j.1365-2672.2005.02552.x.
[20] 丁轲, 倪学勤, 潘康成, 等. 三株乳酸杆菌体外抑菌试验的研究[J]. 饲料工业, 2003, 24(3): 19-21. DOI:10.3969/j.issn.1001-991X.2003.03.007.
[21] 熊涛, 宋苏华, 黄涛, 等. 植物乳杆菌NCU116抑菌性能的研究[J]. 食品与发酵工业, 2012. 38(6): 97-101. DOI:10.13995/j.cnki.11-1802/ ts.2012.06.032.
[22] 宫路路, 龚福明, 李晓然, 等. 豆豉中高产乳酸乳酸菌的筛选及其产酸条件的优化研究[J]. 中国微生态学杂志, 2013, 25(5): 516-520. DOI:10.13381/j.cnki.cjm.2013.05.007.
[23] TULINI F L, WINKELSTROETER L K, de MARTINIS E C P. Identifi cation and evaluation of the probiotic potential of Lactobacillus paraplantarum FT259, a bacteriocinogenic strain isolated from Brazilian semi-hard artisanal cheese[J]. Anaerobe, 2013, 22: 57-63. DOI:10.1016/j.anaerobe.2013.06.006.
[24] DIEULEVEUX V, VANDERPYL D, CHATAUD J, et al. Purifi cation and characterization of anti-Listeria compounds produced by Geotrichum candidum[J]. Applied and Environmental Microbiology, 1998, 64(2): 800-803.
[25] 袁景环, 贡汉生, 孟祥晨. 苯乳酸的抗菌作用及其抗菌机理的初步研究[J]. 食品工业, 2009, 30(5): 14-17.
Antagonistic Mechanism of Lactobacillus plantarum against Foodborne Pathogenic Bacteria
LI Qiangkun, LIU Chenjian, ZHANG Yuanlian, CHANG Linjie, LUO Yiyong, LI Xiaoran*
(Faculty of Life Science and Technology, Kunming University of Science and Technology, Kunming 650500, China)
Abstract:In this study, Oxford cup method was used to screen Lactobacillus plantarum strains with antibacterial effect. The fermentation supernatants from these strains were either treated with protease or adjusted for pH using NaOH before being tested for their ability to inhibit the foodborne pathogenic bacteria Escherichia coli O157:H7 and Staphylococcus aureu. Results showed that organic acid was the major antibacterial compound, yet when adjusted to pH 6.5 the fermentation supernatants had better inhibition ability against E. coli O157:H7. L. plantarum strain QB3-3 showed antibacterial effect for up to 96 h. In addition, the antagonistic mechanism of L. plantarum against foodborne pathogens was due to destruction of the cell structure resulting in morphological changes, leakage of cell contents and fi nally apoptosis as observed via electron microscopy.
Key words:Lactobacillus plantarum; organic acid; antibacterial activity
DOI:10.7506/spkx1002-6630-201611004
中图分类号:Q939.9
文献标志码:A
文章编号:1002-6630(2016)11-0018-06
引文格式:
李强坤, 柳陈坚, 张园莲, 等. 植物乳杆菌对食源性病原菌作用的研究[J]. 食品科学, 2016, 37(11): 18-23. DOI:10.7506/
spkx1002-6630-201611004. http://www.spkx.net.cn LI Qiangkun, LIU Chenjian, ZHANG Yuanlian, et al. Antagonistic mechanism of Lactobacillus plantarum against foodborne pathogenic bacteria[J]. Food Science, 2016, 37(11): 18-23. (in Chinese with English abstract) DOI:10.7506/ spkx1002-6630-201611004. http://www.spkx.net.cn
收稿日期:2015-07-08
基金项目:国家自然科学基金地区科学基金项目(31160309)
作者简介:李强坤(1992—),男,本科,研究方向为生物工程。 E-mail:1764761547@qq.com
*通信作者:李晓然(1984—),女,副教授,博士,研究方向为微生物分子生态学。E-mail:starkeyran@163.com