锦葵色素葡萄糖苷和锦葵色素半乳糖苷抑制内皮细胞氧化应激作用
朱运明
1,2,黄午阳
1,*,闵伟红
2,隋中泉
3,李春阳
1
(1.江苏省农业科学院农产品加工研究所,江苏 南京 210014;2.吉林农业大学食品科学与工程学院,吉林 长春 130118;3.上海交通大学农业与生物学院,上海 200240)
摘 要:目的:研究血管紧张素Ⅱ(angiotensinⅡ,AngⅡ)对人脐静脉内皮细胞(human umbilical vein endothelial cells,HUVEC)中活性氧(reactive oxygen species,ROS)、黄嘌呤氧化酶-1(xanthine oxidase-1,XO-1)、血红素氧化酶-1(heme oxygenase-1,HO-1)含量和超氧化物歧化酶(superoxide dismutase,SOD)活性的影响,确定最佳AngⅡ诱导浓度,探讨不同浓度锦葵色素葡萄糖苷(malvidin-3-glucoside,Mv-3-glc)和锦葵色素半乳糖苷(malvidin-3-galactoside,Mv-3-gal)对AngⅡ诱导的血管内皮细胞氧化应激的抑制作用机理。方法:通过体外培养HUVEC,采用酶联免疫吸附分析法分别测定HUVEC中SOD活力、XO-1和HO-1含量,并采用荧光探针方法检测细胞中ROS水平变化。结果:在HUVEC中,AngⅡ可诱导XO-1的表达,抑制HO-1的表达,AngⅡ在浓度10
-2μmol/L时对HUVEC氧化应激诱导最为显著。不同浓度的Mv-3-glc、Mv-3-gal及其混合物均可显著抑制AngⅡ诱导的内皮细胞氧化应激,通过抑制XO-1蛋白过表达,增加HO-1和SOD表达量,改善细胞抗氧化防御系统,从而减少细胞内ROS水平,达到氧化保护作用,且Mv-3-glc比Mv-3-gal抗氧化活性更强。结论:锦葵色素葡萄糖苷和锦葵色素半乳糖苷具有抑制内皮细胞氧化应激的作用。
关键词:锦葵色素葡萄糖苷;锦葵色素半乳糖苷;内皮细胞;血管紧张素Ⅱ;活性氧;氧化应激
朱运明, 黄午阳, 闵伟红, 等. 锦葵色素葡萄糖苷和锦葵色素半乳糖苷抑制内皮细胞氧化应激作用[J]. 食品科学, 2016, 37(17): 201-206. DOI:10.7506/spkx1002-6630-201617034. http://www.spkx.net.cn
ZHU Yunming, HUANG Wuyang, MIN Weihong, et al. Effects of malvidin glycosides on AngⅡ-induced oxidative stress in endothelial cells[J]. Food Science, 2016, 37(17): 201-206. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201617034. http://www.spkx.net.cn
心血管疾病是危害人类健康的头号杀手,而80%~90%心血管疾病患者都伴随着高血压
[1]。肾素-血管紧张素-醛固酮系统(renin-angiotensin-aldosterone system,RAAS)激活是高血压发病机制的主要环节之一
[2]。近来研究发现血管紧张素Ⅱ(angiotensinⅡ,AngⅡ)作为RAAS系统最重要的效应分子,它可以产生收缩血管、增加心肌收缩力、促细胞增殖、诱导细胞产生过量的活性氧(reactive oxygen species,ROS)引发氧化应激反应
[3]。
氧化应激损害细胞结构与功能从而导致心血管疾病
[4]。ROS是体内氧化应激的主要来源,大量ROS累积会造成细胞损伤
[5]。血管内皮细胞参与产生ROS的酶有多种,如黄嘌呤氧化酶-1(xanthine oxidase-1,XO-1)、烟酰胺腺嘌呤二核苷磷酸(nicotinamide adenine dinucleotide 2’-phosphate reduced tetrasodium salt,NADPH)等
[6]。XO-1催化由黄嘌呤形成尿酸以及次黄嘌呤形成黄嘌呤的氧化过程,是内皮细胞中过氧化物的主要来源
[7-8]。AngⅡ可激活内皮细胞XO-1,促进细胞中氧化应激反应,引起内皮细胞功能性障碍,从而加速心血管等疾病进程
[9-10]。血红素氧化酶-1(heme oxygenase-1,HO-1)催化血红素降解,产生一氧化碳、游离铁和胆绿素,表现出很强的抗氧化效果,延缓动脉粥样硬化
[11-13]。超氧化物歧化酶(superoxide dismutase,SOD)催化有毒的超氧化物自由基转变为普通分子氧和过氧化氢
[14]。HO-1和SOD都参与细胞抗氧化防御机制。目前AngⅡ对人脐静脉内皮细胞(human umbilical vein endothelial cells,HUVEC)中XO-1、HO-1、SOD表达情况的影响尚不明确。
抗氧化剂能够清除自由基,削弱氧化应激和减轻ROS相关的有害影响,有益于防治各种与自由基相关的疾病。花青素被证实是自然界最有效的水溶性抗氧化剂,有增强心脏功能、抗心血管疾病、抗衰老、抗癌及抗突变等多种生理活性功能
[15]。锦葵色素是兔眼蓝莓(Vaccinium ashei)中含量最高的花青素,主要以锦葵色素葡萄糖苷(malvidin-3-glucoside,Mv-3-glc)和锦葵色素半乳糖苷(malvidin-3-galactoside,Mv-3-gal)形式存在。前期研究发现锦葵色素及其糖苷具有良好的体外抗氧化、抗炎、血管紧张素转换酶(angiotensin I-converting enzyme,ACE)抑制作用
[16-18]。因此,本研究以HUVEC模型,利用荧光显微镜检测细胞ROS水平,采用酶联免疫吸附分析(enzyme linked immunesorbent assay,ELISA)法从蛋白水平上检测XO-1、HO-1、SOD的表达,阐明AngⅡ引起内皮细胞氧化应激的作用,进一步探讨Mv-3-glc、Mv-3-gal的抗氧化作用机制。
1 材料与方法
1.1 材料与试剂
HUVEC 上海中乔新舟生物科技有限公司;AngⅡ、氯化Mv-3-glc、氯化Mv-3-gal 美国Sigma公司;改良Eagle培养基(Dulbecco’s modified Eagle medium,DMEM)高糖培养液、乙二胺四乙酸(ethylene diamine tetraacetic acid,EDTA)胰酶及胎牛血清美国Gibco公司;ROS检测试剂盒 碧云天生物技术研究所;XO-1、HO-1、SOD-ELISA试剂盒 上海博谷生物科技有限公司。
1.2 仪器与设备
CO
2细胞培养箱 美国热电公司;LB941微孔板式多功能分析仪 德国Berthold Technology公司;荧光倒置显微镜IX53 日本Olympus公司。
1.3 方法
1.3.1 HUVEC的培养
取出液氮中保存的HUVEC,用含有质量分数10%胎牛血清、2%青链霉素的DMEM高糖培养液,在37 ℃、5% CO
2培养箱中复苏细胞,再用质量分数0.25% EDTA胰酶进行消化,细胞计数、传代、冻存。选择生长良好的3~7 代细胞用于实验。
1.3.2 细胞分组处理
HUVEC以1×10
5个/mL的密度接种于6 孔板,细胞生长融合成单层(长满单层80%~90%)后用于实验。实验前用无血清DMEM饥饿培养4 h,使各孔中细胞状态达到一致。
为确定最佳刺激浓度,采用不同浓度AngⅡ对细胞进行刺激。实验分6 组(每孔1 mL细胞悬液加入试剂10 μL,下同):空白组(二甲基亚砜(dimethyl sulfoxide,DMSO))、浓度分别为10
-3、10
-2、10
-1、1、10 μmol/L的 AngⅡ刺激组,AngⅡ刺激24 h后,测定各项指标变化。
为考察锦葵色素糖苷对HUVEC的抗氧化作用,样品设立分组:空白组(DMSO)、AngⅡ刺激组(10
-2μmol/L)、终浓度为1、5、10 μmol/L Mv-3-glc+AngⅡ(10
-2μmol/L)实验组;终浓度为1、5、10 μmol/L Mv-3-gal+ AngⅡ(10
-2μmol/L)实验组;终浓度为1、5、10 μmol/L Mv-3-glc与Mv-3-gal的混合液(二者物质的量比为1∶1)+AngⅡ(10
-2μmol/L)实验组。预先加入不同浓度的样品处理18 h,再加入AngⅡ培养6 h,测定各项指标变化。
1.3.3 ELISA测定细胞上清液中SOD活力、HO-1、XO-1蛋白含量
细胞经不同分组处理后,无菌收集培养液上清液,1 000×g离心15 min,收集上清液,用于ELISA检测。ELISA操作严格按照试剂盒说明书进行。上清液按一定比例稀释(空白组加等体积无血清培养液),在酶标仪450 nm波长处测定吸光度(A
450
nm)。根据样品的吸光度在标准曲线上找出对应的浓度,乘以稀释度计算样品的最终浓度,测定出SOD活力、HO-1、XO-1蛋白含量,其相对活力或含量是相对空白组所得。
1.3.4 HUVEC中ROS含量水平的测定
处理后的细胞弃去上清液,在无菌条件下装载2’,7’-二氯荧光黄双乙酸盐(2’,7’-dichlorofluorescin diacetate,DCFH-DA)探针(终浓度10 μmol/L),37 ℃细胞培养箱内孵育20 min,用无血清DMEM培养液或无菌磷酸缓冲盐溶液(phosphate buffer saline,PBS)洗涤细胞3 次,以充分去除未进入细胞内的DCFH-DA。在暗室中用荧光显微镜进行荧光拍照观察各组细胞中ROS含量水平,并用荧光酶标仪(激发波长485 nm,发射波长535 nm)检测荧光强度,以荧光强度表示细胞中ROS含量,而ROS相对含量则是相对于空白组荧光强度。操作严格按照试剂盒说明书进行。
1.4 数据统计分析
数据以SPSS 18.0统计软件进行统计分析。所有结果以±s表示,所有样品测定3 次。多组均数间采用单因素方差分析,两两之间比较采用Dunnett-t检验。当P<0.05时表示存在显著性差异。
2 结果与分析
2.1 AngⅡ最佳刺激浓度的确定
由表1可知,正常细胞中ROS水平较低,荧光强度较弱。AngⅡ刺激HUVEC后,胞内ROS含量产生不同程度的增加。随着AngⅡ浓度的增大,细胞内ROS含量先增大后减小,当AngⅡ浓度为10
-2μmol/L时,胞内ROS含量最高,荧光强度达到2 653.33±96.65(P<0.01)。低浓度的AngⅡ(10
-3μmol/L)对细胞刺激不够,而高浓度的AngⅡ(1、10 μmol/L)对细胞生长产生了抑制作用,使得ROS含量降低。
表1 不同浓度AngⅡ对HUVEC细胞中ROS及上清液中XO-1和HO-1含量的影响 (x± s,, n = 3)
Table 1 Effect of different concentration AngⅡon ROS in HUVECs, and XO-1 and HO-1 production in supernatants (x± s,, n = 3)
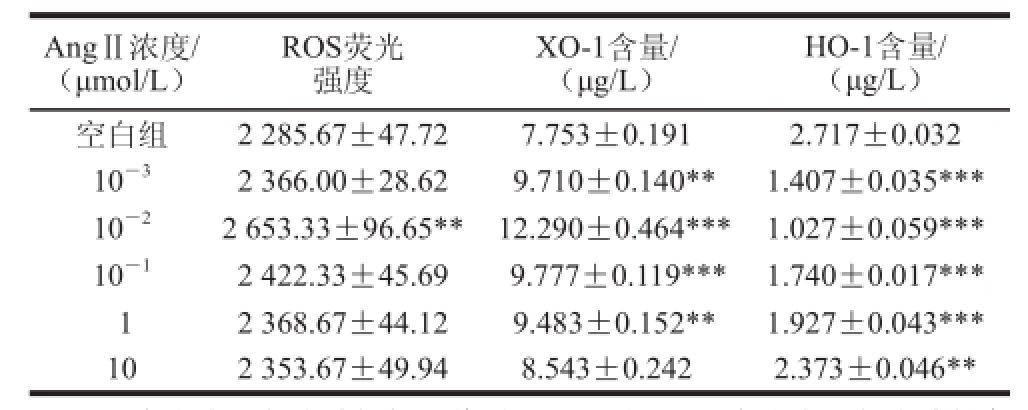
注:**. 与空白组相比差异极显著(P<0.01);***. 与空白组相比差异高度显著(P<0.001)。
HO-1含量/(μg/L)AngⅡ浓度/(μmol/L)ROS荧光强度XO-1含量/(μg/L)空白组2 285.67±47.727.753±0.1912.717±0.032 10
-32 366.00±28.62 9.710±0.140**1.407±0.035*** 10
-22 653.33±96.65** 12.290±0.464***1.027±0.059*** 10
-12 422.33±45.69 9.777±0.119***1.740±0.017*** 12 368.67±44.12 9.483±0.152**1.927±0.043*** 102 353.67±49.948.543±0.2422.373±0.046**
由表1还可知,空白组细胞表达少量的XO-1蛋白((7.753±0.191) μg/L),而HO-1蛋白表达量相对较高((2.717±0.032) μg/L),当不同浓度AngⅡ刺激后,细胞上清液中XO-1蛋白表达量增加,而HO-1蛋白表达量减少。随着AngⅡ浓度增大,XO-1和HO-1蛋白水平的变化先增大后减小,这表明高浓度的AngⅡ对细胞生长有抑制作用。当AngⅡ浓度为10
-2μmol/L时,XO-1蛋白的表达量最高达到(12.290±0.464) μg/L,为空白组的1.59 倍 (P<0.001);同时,HO-1蛋白表达量最低达到(1.027±0.059) μg/L,为空白组的0.38 倍(P<0.001),具有统计学意义。高浓度的AngⅡ(10 μmol/L)对HUVEC中XO-1蛋白表达影响不显著(P>0.05),其表达量接近空白组。结果表明:10
-2μmol/L的AngⅡ对HUVEC中XO-1和HO-1蛋白表达影响极其显著,对ROS水平影响也最显著,因此确定AngⅡ最佳刺激浓度为10
-2μmol/L。
2.2 锦葵色素糖苷对HUVEC中ROS相对含量的影响
AngⅡ(10
-2μmol/L)刺激HUVEC时,内皮细胞中超氧化物相对含量增加。锦葵色素糖苷预处理HUVEC,可以保护细胞减少氧化应 激反应,使ROS相对含量降 低。随着样品浓度的增大,细胞内ROS相对含量减小。由图1可知,1、5、10 μmol/L的Mv-3-glc处理细胞,对AngⅡ刺激后增加的ROS相对含量分别抑制了69%、103%、115%;Mv-3-gal处理细胞,对AngⅡ刺激增加的ROS相对含量抑制率均超过了100%,ROS相对含量为空白组的0.908、0.851、0.877 倍;Mv-3-glc+Mv-3-gal处理细胞,ROS相对含量与AngⅡ刺激组相比也显著减少,ROS相对含量为空白组的0.946、0.898、0.903 倍。因此,锦葵色素糖苷对HUVEC中ROS相对含量下降影响高度显著(P<0.001),可以保护HUVEC免受AngⅡ引起的氧化应激反应。

图1 不同浓度锦葵色素糖苷对AngⅡ刺激的HUVEC中ROS水平的影响
Fig. 1 Effects of Mv-3-glc, Mv-3-gal, and their mixture on the level of AngⅡ-induced ROS in HUVECs
2.3 锦葵色素糖苷对HUVEC中XO-1相对含量的影响
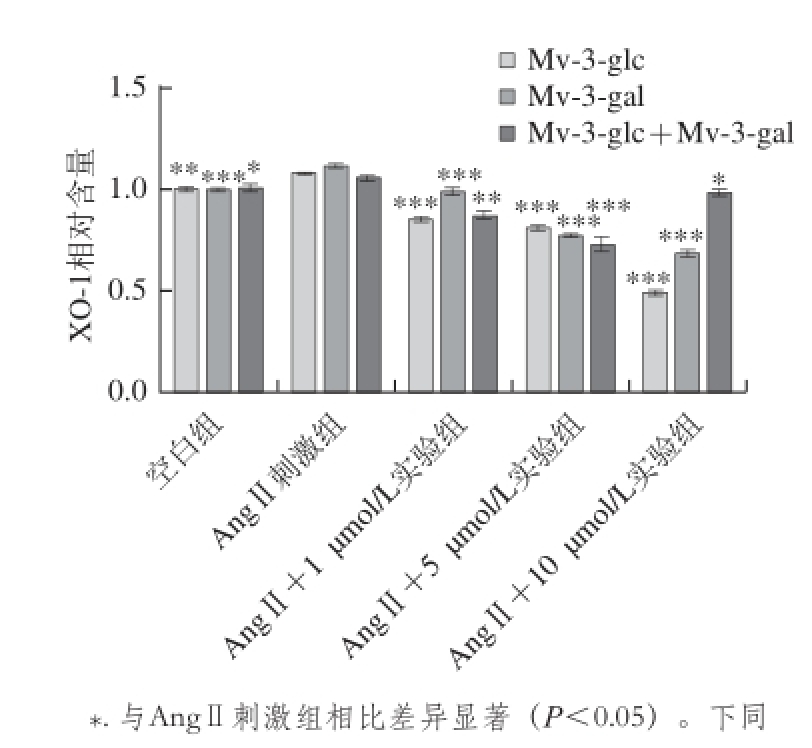
图2 不同浓度锦葵色素糖苷对AngⅡ诱导的HUVEC中XO-1水平的影响
Fig. 2 Effects of Mv-3-glc, Mv-3-gal, and their mixture on the level of AngⅡ-induced XO-1 in HUVECs
由图2可知,AngⅡ(10
-2μmol/L)刺激HUVEC时,内皮细胞上清液中XO-1相对含量显著增加。锦葵色素糖苷预处理HUVEC,可以显著抑制XO-1相对含量增加。对于Mv-3-glc和Mv-3-gal处理实验组,随着二者浓度的增大,细胞内XO-1相对含量呈浓度依赖性减小,1、5、10 μmol/L的样品预处理细胞,XO-1相对含量比AngⅡ刺激组显著降低,分别为空白组的0.85、0.81、0.49 倍和0.98、0.77、0.68 倍。Mv-3-glc比Mv-3-gal抗氧化活性更强,但两者并没有协同效应。两者的混合液Mv-3-glc+Mv-3-gal预处理细胞,对HUVEC细胞中XO-1相对含量的影响随浓度的增加先增强后减弱,相对含量分别为空白组的0.86、0.73、0.98 倍。因此,锦葵色素糖苷对HUVEC细胞中XO-1相对含量的影响显著(P<0.05),能完全保护HUVEC免受AngⅡ引起的氧化损伤。
2.4 锦葵色素糖苷对HUVEC中HO-1相对含量的影响
AngⅡ(10
-2μmol/L)刺激HUVEC时,内皮细胞上清液中HO-1相对含量显著降低。锦葵色素糖苷预处理HUVEC,HO-1相对含量比AngⅡ刺激组要高,但比空白组低。随着样品浓度的增大,细胞内HO-1相对含量先增加后减小,1、5、10 μmol/L的Mv-3-glc、Mv-3-gal、Mv-3-glc+Mv-3-gal预处理细胞,HO-1相对含量分别为AngⅡ刺激组的1.55、3.27、1.68 倍,1.56、2.48、2.29 倍,1.55、3.28、1.68 倍。在浓度为5 μmol/L时,抗氧化活性达到最强,抑制率分别为81.9%、57.6%、82.3%。由图3可知,锦葵色素糖苷对HUVEC细胞中HO-1相对含量的影响显著(P<0.05),可以保护HUVEC,减弱AngⅡ引起的HO-1相对含量降低,起到抗氧化作用。在低浓度情况下,Mv-3-glc相对Mv-3-gal有更好的抗氧化活性,两者的混合液活性高于Mv-3-gal。
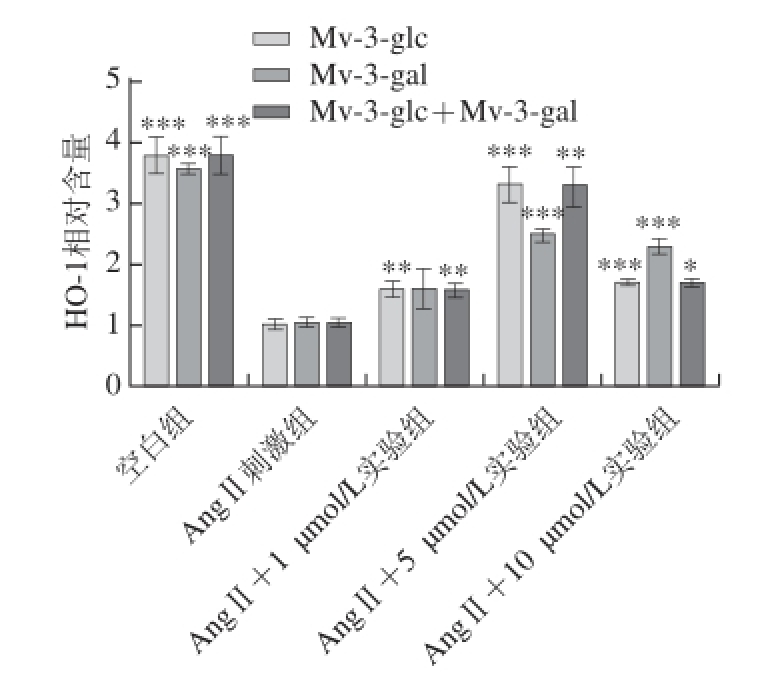
图3 不同浓度锦葵色素糖苷对AngⅡ诱导的HUVEC中HO-1水平的影响
Fig. 3 Effects of Mv-3-glc, Mv-3-gal, and their mixture on Ang Ⅱ-induced HO-1 in HUVECs
2.5 锦葵色素糖苷对HUVEC中SOD相对活力的影响
AngⅡ(10
-2μmol/L)刺激HUVEC时,内皮细胞上清中SOD相对活力略有降低。锦葵色素糖苷预处理HUVEC,SOD相对活力升高。Mv-3-glc的抗氧化作用极其显著,由实验得到1、5、10 μmol/L预处理细胞SOD活力高达(652.59±28.92)、(1062.79±12.19)、(608.32±54.56) U/mg,由图4可知,分别为空白组的2.98、4.85、2.77 倍(AngⅡ组为空白组的0.89 倍);Mv-3-gal及Mv-3-glc+Mv-3-gal混合液预处理细胞,SOD相对活力分别为空白组的1.40、2.06、1.43 倍(AngⅡ组为空白组的0.91 倍)和1.08、1.83、1.54 倍(AngⅡ组为空白组的0.94 倍)。综上所述,锦葵色素糖苷在HUVEC细胞中是通过增加SOD活性和HO-1含量、降低XO-1含量等多个途径减少细胞内ROS水平,显著提高其抗氧化活性,可以保护HUVEC免受AngⅡ引起的氧化应激。

图4 不同浓度锦葵色素糖苷对AngⅡ诱导的HUVEC中SOD水平的影响
Fig. 4 Effects of Mv-3-glc, Mv-3-gal, and their mixture on Ang II-induced SOD in HUVECs
3 讨 论
高血压主要是遗传易感性和环境因素相互作用的结果,引起该疾病的因素很多,如血胆脂醇过多、糖尿病和吸烟等,都会增加血管中ROS的含量而导致内皮细胞功能障碍
[19]。此外,氧化应激可导致细胞中ACE过量表达,造成细胞氧化损伤,加速血管疾病的进程
[20]。研究表明,AngⅡ可刺激内皮细胞分泌单核细胞趋化蛋白-1和细胞间黏附分子-1,参与动脉粥样硬化
[21-22]。本实验结果表明AngⅡ还可以激活XO-1和抑制HO-1表达,促进氧化应激反应导致细胞中ROS含量升高,对血管内皮细胞造成损伤,而Mv-3-gal及Mv-3-glc能够保护内皮细胞,显著抑制氧化应激反应,减少细胞ROS和XO-1含量,改善细胞抗氧化防御系统,增加HO-1和SOD表达。
本实验以HUVEC为研究对象,以不同浓度AngⅡ(10
-3、10
-2、10
-1、1、10 μmol/L)对HUVEC中产生过氧化物的XO-1和表现抗氧化功能的HO-1表达量的影响,反映AngⅡ在血管内皮细胞氧化应激中的功能作用。结果表明不同浓度的AngⅡ与HUVEC共育24 h后,XO-1和HO-1表达量变化显著,细胞上清液中XO-1含量增加,HO-1含量减少;此外细胞中ROS水平也相应提高。当AngⅡ浓度为10
-2μmol/L时,XO-1的表达量达到最高,而HO-1的表达量达到最低,均具有统计学意义(P<0.001)。当AngⅡ浓度进一步增加时,高浓度AngⅡ会抑制HUVEC的生长,XO-1和HO-1的含量反而减少。XO-1在HUVEC中表达会引起胞内ROS含量的升高,HO-1在HUVEC中表达会引起胞内ROS含量的降低。实验中,从胞内ROS来看,当AngⅡ刺激HUVEC时,ROS含量是先增大后减小,这与XO-1表达量关系一致,而与HO-1表达量关系相反,尤其当AngⅡ浓度为10
-2μmol/L时,胞内ROS含量达到最高,与胞内XO-1和HO-1表达一一对应,也具有统计学意义(P<0.01)。然而AngⅡ对细胞中SOD活性没有显著影响。
不同浓度的Mv-3-gal及Mv-3-glc及其混合物预处理HUVEC,细胞上清液中XO-1含量减少,HO-1含量和SOD活性增加;此外细胞中ROS水平也相应降低,这揭示了锦葵色素及其糖苷在细胞内的抗氧化机制。与体外分光光度法测定抗氧化活性不同,在细胞内测定抗氧化作用,化合物并没有显示出良好的线性量效关系,这是由于细胞本身存在个体差异,且影响细胞生长和蛋白生成量的因素众多,而高剂量药物往往会制约细胞生长造成蛋白含量降低。一般情况下,糖基化结构减少自由基结合位点会降低花青素的抗氧化活性
[23],然而Kähkönen等
[24]发现糖基化模式对花青素抗氧化功效的影响取决于配基结构以及所测定的抗氧化分析模型,不同的糖基化环境可能减弱抗氧化活性,也可能增强抗氧化活性。实验表明大多数情况下Mv-3-glc比Mv-3-gal抗氧化活性更强,此外Mv-3-gal及Mv-3-glc有时表现出一定的协同效应,然而两者之间的关系还需进一步的实验分析来证明。
本研究从分子基础上研究Mv-3-gal及Mv-3-glc对AngⅡ诱导的内皮细胞氧化应激的抑制作用机制。抗氧化功能与保护心血管功能有关,结果表明Mv-3-glc、Mv-3-gal及两者混合物可抑制由AngⅡ诱导的内皮细胞ROS和XO-1过表达,同时增加内源性细胞抗氧化保护物HO-1含量和SOD活性,降低细胞氧化应激,保护血管内皮细胞免受损伤。
综上所述,花青素成分Mv-3-gal及Mv-3-glc具有保护血管内皮细胞免受氧化损伤作用,可作为一种潜在的抗氧化剂,对防治心血管疾病起到积极作用,具有良好的应用价值。
参考文献:
[1] WANG J G, LI Y. Characteristics of hypertension in the Chinese population[J]. Current Hypertension Reports, 2012, 14(5): 410-415. DOI:10.1007/s11906-012-0288-1.
[2] HSUEH W A, WYNE K. Renin-angiotensin-aldostero ne system in diabetes and hypertension[J]. The Journal of Clinical Hypertension, 2011, 13(4): 224-237. DOI:10.1111/j.1751-7176.2011.00449.x.
[3] BELOVA L A. Angiotensin Ⅱ-generating enzymes[J]. Biochemistry (Moscow), 2000, 65(12): 1337-1345. DOI:10.1023/A:1002848402911. [4] PIALOUX V, MOUNIER R. Hypoxia-induced oxidative stress in health disorders[J]. Oxidative Medicine and Cellular Longevity, 2012: 940121. DOI:10.1155/2012/940121.
[5] ZAMORA-ROS R, FEDIRKO V, TRICHOPOULOU A, et al. Dietary flavonoid, lignan and antioxidant capacity and risk of hepatocellular carcinoma in European prosective investigation into cancer and nutrition study[J]. International Journal of Cancer, 2013, 133: 2429-2443. DOI:10.1002/ijc.28257.
[6] GRIENDLING K K, FITZGERALD G A. Oxidative stress and cardiovascular injury part Ⅰ: basic mechanisms and in vivo monitoring of ROS[J]. Circulation, 2003, 108(16): 1912-1916. DOI:10.1161/01. CIR.0000093660.86242.BB.
[7] NOMURA J, BUSSO N, IVES A, et al. Xanthine oxidase inhibition by febuxostat attenuates experimental atherosclerosis in mice[J]. Scientifi c Reports, 2014, 4: 4554. DOI:10.1038/srep04554.
[8] LANDMESSER U, SPIEKERMANN S, PREUSS C, et al. Angiotensin Ⅱinduces endothelial xanthine oxidase activation role for endothelial dysfunction in patients with coronary disease[J]. Arteriosclerosis, Thrombosis, and Vascular Biology, 2007, 27(4): 943-948. DOI:10.1161/01.ATV.0000258415.32883.bf.
[9] LANDMESSER U, SPIEKERMANN S, DIKALOV S, et al. Vascular oxidative stress and endothelial dysfunction in patients with chronic heart failure role of xanthine-oxidase and extracellular superoxide dismutase[J]. Circulation, 2002, 106(24): 3073-3078. DOI:10.1161/01. CIR.0000041431.57222.AF.
[10] LANG D, MOSFER S I, SHAKESBY A, et al. Coronary microvascular endothelial cell redox state in left ventricular hypertrophy the role of angiotensin Ⅱ[J]. Circulation Research, 2000, 86(4): 463-469. DOI:10.1161/01.RES.86.4.463.
[11] ERDMANN K, GROSSER N, SCHRÖDER H. L-Methionine reduces oxidant stress in endothelial cells: role of heme oxygenase-1, ferritin, and nitric oxide[J]. The AAPS Journal, 2005, 7(1): E195-E200. DOI:10.1208/aapsj070118.
[12] GROSSER N, OBERLE S, BERNDT G, et al. Antioxidant action of L-alanine: heme oxygenase-1 and ferritin as possible mediators[J]. Biochemical and Biophysical Research Communications, 2004, 314(2): 351-355.
[13] ERDMANN K, GROSSER N, SCHIPPOREIT K, et al. The ACE inhibitory dipeptide Met-Tyr diminishes free radical formation in human endothelial cells via induction of heme oxygenase-1 and ferritin[J]. The Journal of Nutrition, 2006, 136(8): 2148-2152.
[14] RAYCHAUDHURI S S, DENG X W. The role of superoxide dismutase in combating oxidative stress in higher plants[J]. The Botanical Review, 2008, 66(1): 89-98. DOI:10.1007/BF0285778.
[15] POJER E, MATTIVI F, JOHNSON D, et al. The case for anthocyanin consumption to promote human health: a review[J]. Comprehensive Reviews in Food Science and Food Safety, 2013, 12(5): 483-508. DOI:10.1111/1541-4337.12024.
[16] POP R, STEFANUT M N, CATA A, et al. Ab initio study regarding the evaluation of the antioxidant character of cyanidin, delphinidin and malvidin[J]. Central European Journal of Chemistry, 2012, 10(1): 180-186. DOI:10.2478/s11532-011-0128-1.
[17] LEE C, HAN D, KIM B, et al. Antioxidant and anti-hypertensive activity of anthocyanin-rich extracts from hulless pigmented barley cultivars[J]. International Journal of Food Science and Technology, 2013, 48(5): 984-991. DOI:10.1111/ijfs.12050.
[18] HUANG W Y, WANG J, LIU Y M, et al. Inhibitory effect of malvidin on TNF-α-induced inflammatory response in endothelial cells[J]. European Journal of Pharmacology, 2014, 723: 67-72. DOI:10.1016/ j.ejphar.2013.11.041.
[19] CALHOUN D A, JONES D, TEXTOR S, et al. Resistant hypertension: diagnosis, evaluation, and treatment. A scientifi c statement from the American Heart Association Professional Education Committee of the Council for High Blood Pressure Research[J]. Hypertension, 2008, 51(6): 1403-1419. DOI:10.1161/CIRCULATIONAHA.108.189141.
[20] HIGASHI Y, NOMA K, YOSHIZUMI M, et al. Endothelial function and oxidative stress in cardiovascular diseases[J]. Circulation Journal, 2009, 73(3): 411-418. DOI:10.1253/circj.CJ-08-1102.
[21] PASTORE L, TESSITORE A, MARTINOTTI S, et al. Angiotensin Ⅱstimulates intercellular adhesion molecule-1 (ICAM-1) expression by human vascular endothelial cells and increases soluble ICAM-1 release in vivo[J]. Circulation, 1999, 100(15): 1646-1652.
[22] WHITE C R, DARLEY-USMAR V, BERRINGTON W R, et al. Circulating plasma xanthine oxidase contributes to vascular dysfunction in hypercholesterolemic rabbits[J]. Proceedings of the National Academy of Sciences, 1996, 93(16): 8745-8749.
[23] MIGUEL M G. Anthocyanins: antioxidant and/or anti-inflammatory activities[J]. Journal of Applied Pharmaceutical Science, 2010, 1: 7-15.
[24] KÄHKÖNEN M P, HEINONEN M. Antioxidant activity of anthocyanins and their aglycons[J]. Journal of Agricultural and Food Chemistry, 2003, 51: 628-633. DOI:10.1021/jf025551i.
Effects of Malvidin Glycosides on Ang II-Induced Oxidative Stress in Endothelial Cells
ZHU Yunming
1,2, HUANG Wuyang
1,*, MIN Weihong
2, SUI Zhongquan
3, LI Chunyang
1
(1. Institute of Farm Product Processing, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China; 2. College of Food Science and Engineering, Jilin Agricultural University, Changchun 130118, China; 3. School of Agriculture and Biology, Shanghai Jiao Tong University, Shanghai 200240, China)
Abstract:The objective of this research was to investigate the antioxidant role in endothelial cells. The influence of angiotensin II (Ang II) on the expression of xanthine oxidase-1 (XO-1) and heme oxygenase-1 (HO-1) in human umbilical vein endothelial cells (HUVECs), and the changes in reactive oxygen species (ROS), superoxide dismutase (SOD), XO-1, and HO-1 caused by two malvidin glycosides malvidin-3-glucoside (Mv-3-glc) and malvidin-3-galactoside (Mv-3-gal) were investigated. The levels of XO-1, HO-1, and SOD were evaluated by enzyme linked inmunosorbent assay (ELISA) kits. ROS level was detected under a fl uorescence microplate. Compared with the control group, the expression of XO-1 protein was signifi cantly increased and the expression of HO-1 protein was signifi cantly decreased in the 10
-2μmol/L Ang II-treated group. Different concentrations of Mv-3-glc and Mv-3-gal as well as their mixture greatly decreased the levels of ROS and XO-1, but increased the levels of SOD and HO-1. Glycosides improved the antioxidant capacity of malvidin to a great extent. The antioxidant activity of Mv-3-glc was more pronounced than that of Mv-3-gal. Variation in malvidin glycosides had a signifi cant impact on Ang II-induced oxidative stress to different extents. Therefore, malvidin glycosides can protect cells from oxidative deterioration and thus used to prevent diseases associated with oxidative stress.
Key words:malvidin-3-glucoside; malvidin-3-galactoside; endothelial cells; angiotensin II (Ang II); reactive oxygen species (ROS); oxidative stress
DOI:10.7506/spkx1002-6630-201617034
中图分类号:R329.25;R332
文献标志码:A
文章编号:1002-6630(2016)17-0201-06
收稿日期:2015-09-28
基金项目:国家自然科学基金青年科学基金项目(NSFC31101264)
作者简介:朱运明(1988—),男,硕士研究生,研究方向为发酵微生物的选育与代谢调控。E-mail:zhuming.1988.happy@163.com
*通信作者:黄午阳(1979—),女,副研究员,博士,研究方向为功能食品与活性物质。E-mail:wuyanghuang@hotmail.com
引文格式: