表1 鸡肉肠感官评分标准
Table1 Criteria for sense evaluation of chicken sausage

白青云,许 倩,李玉玉,李 雯,殷依琳,赵 立
(淮阴工学院生命科学与食品工程学院,江苏 淮安 223003)
摘 要:采用人工接种乳酸菌的方法,对发酵鸡肉肠中γ-氨基丁酸(γ-amino butyric acid,GABA)进行富集。首先鸡肉肠中分别添加不同来源的乳酸菌,筛选GABA富集的最佳发酵菌种;然后研究外源添加物L-谷氨酸(L-Glu)、VB6和CaCl2对鸡肉肠中GABA含量的影响,并采用Box-Behnken试验设计优化添加量。结果表明,3 种不同来 源的乳酸菌发酵鸡肉肠,其中酸奶乳酸菌与泡菜乳酸菌产GABA的能力较弱,均低于10 mg/100 g,耐久肠球菌产GABA能力最强,GABA含量达到62.14 mg/100 g,显著高于其他两种菌(P<0.05);Box-Behnken设计得到发酵鸡肉肠富集GABA的最优外源添加物添加量为L-Glu 7.75 mg/100 g、VB66.73 mg/100 g、CaCl28.35 mg/100 g,在此条件下鸡肉肠中GABA含量为68.32 mg/100 g,是未添加外源物含量的1.10 倍,比普通鸡肉肠约提高10 倍。方差分析表明,所建的回归模型能够很好地预测鸡肉肠中GABA含量的变化。其中,3 种外源添加物的添加量均极显著影响鸡肉肠中GABA含量(P<0.01),L-Glu和VB6添加量的交互作用以及L-Glu和CaCl2添加量的交互作用均显著影响鸡肉肠GABA含量(P<0.05)。
关键词:发酵鸡肉肠;γ-氨基丁酸;乳酸菌;条件优化
发酵鸡肉肠是以鸡肉和猪脂肪为主要原料,结合西式发酵香肠工艺而制成的一种方便食品,因此同时具有鸡肉和发酵香肠的特点与营养价值[1]。香肠发酵成熟后,鸡肉中的蛋白质分解为氨基酸,促进人体消化吸收;同时增加了必需氨基酸及维生素等物质,加强了肉制品的保健性和营养性;另外也可进一步生成大量香味,产生特殊风味[2-3]。肉制品中人工接种乳酸菌发酵后,产生大量有益微生物,不仅抑制了肉制品腐败,延长了保质期,而且有利于降低亚硝胺的含量,减少对人体的伤害,提高了产品的安全性[4-5]。
γ-氨基丁酸(γ-amino butyric acid,GABA)是哺乳动物大脑和脊髓中重要的抑制性神经传递物质,具有降低血压、调节心跳速率,缓解疼痛和紧张等功能[6]。近年来研究表明,GABA能够改善人体免疫功能并通过清除自由基保护免疫细胞[7]。由于GABA的多重功能,目前关于功能食品富集GABA的方法屡见报道。GABA富集方法有植物富集法和微生物发酵法[8],微生物发酵法生产富含GABA的食品成为近年来研究的热点,而安全性较高的乳酸菌成为用于富集GABA的首选菌种[9],如许建军等[10]筛选到高产GABA的乳酸菌,梁金钟等[11]采用诱变技术使乳酸菌产GABA量提高1.979 倍。GABA由L-谷氨酸(L-Glu)在谷氨酸脱羧酶(glutamate decarboxylase,GAD)催化下脱羧而成,GAD活性受多种因素的影响,添加底物或酶激活剂均可促进GABA的积累[12-13]。L-Glu作为生产GABA的前体物质,对GABA的积累具有重要影响。在不添加外源L-Glu的条件下,菌体自身合成的L-Glu仅有少量转化为GABA,添加外源L-Glu可使GABA代谢支路中碳流量增加,并可调节GAD活性,进而获得大量的GABA[14]。GAD含有CaM结合区,GAD活性受H+和Ca2+浓度的调控[15],除此外,GAD活力还受GAD辅酶——磷酸吡哆醛(pyridoxal phosphate,PLP)的影响,一定质量浓度的PLP提高GAD活性,促进GABA积累[16]。VB6是PLP的前体物质,在生命体中可转化为PLP,有激活GAD达到富集GABA的作用[17],VB6较PLP廉价,在生产上更具现实意义。
鸡肉肠中蛋白含量较高,一般在11%左右[18],是很好的蛋白食品。鸡肉中的蛋白质在发酵过程中会发生降解作用,生成小分子可溶性蛋白质、游离氨基酸、多肽等[19]。本研究采用人工接种乳酸菌发酵鸡肉,首先筛选高产GABA的乳酸菌种,然后研究外源添加物对鸡肉肠中GABA的影响,并采用响应面法优化富集条件,旨在为生产富含GABA的发酵鸡肉肠提供理论依据。
1.1 材料、菌种与试剂
鸡脯肉、猪脂肪、肠衣等购于淮安市当地农贸市场;食盐、蔗糖、调味料等购于淮安市商联超市。
酸奶乳酸菌(保加利亚乳杆菌和嗜热链球菌)购于川秀旗舰店;泡菜乳酸菌由本校生物工程实验室自发酵泡菜中分离制得;耐久肠球菌Enterococcus durans(编号20379)购于中国工业微生物菌种保藏中心。
GABA标准品和其他试剂均购自国药化学试剂有限公司。
1.2 仪器与设备
HH-6型数显恒温水浴锅 常州国华电器有限公司;TDL-40B型离心机 上海安亭仪器厂;722型可见分光光度计 上海精密科学仪器有限公司;PHSJ-5型pH计上海雷磁电化学分析仪器公司;SPX-250B-Z型生化培养箱 上海博迅实业有限公司医疗设备厂;YM-100型立式压力蒸汽灭菌锅、SW-CJ-1F超净工作台 上海菁华科技仪器有限公司。
1.3 方法
1.3.1 发酵鸡肉肠制作工艺流程[20]
原料肉预处理→混料→斩拌→接种→搅拌→灌肠→漂洗与打孔→发酵→成熟→烘干→成品
1.3.2 不同来源乳酸菌对发酵鸡肉肠中GABA含量的影响
将酸奶乳酸菌、泡菜乳酸菌、耐久肠球菌用培养基扩大培养后离心,得到菌体,然后用0.85%的生理盐水溶解,配成0.50%、0.75%、1.0%、1.25%、1.50%的溶液,人工接种到鸡肉肠中,在30 ℃发酵48 h,测定发酵鸡肉肠中GABA含量,筛选高产GABA的最佳菌种。
1.3.3 外源添加物对发酵鸡肉肠GABA含量和感官评分的影响
采用1.3.2节筛选的最佳菌种按最高产GABA的接种量接种鸡肉肠,对影响鸡肉肠GABA富集的外源添加物L-Glu、VB6和CaCl2的添加量进行研究。各组分的添加范围分别为:L-Glu 3~12 mg/100 g、VB63~12 mg/100 g、CaCl25~20 mg/100 g。在30 ℃发酵48 h,测定鸡肉肠中GABA含量。为了评价产品品质,同时对产品进行感官评分。对影响鸡肉肠GABA含量的添加物浓度采用Box-Behnken设计进行优化,以GABA含量作为考察值。
1.3.4 指标测定
1.3.4.1 GABA含量测定
将一定量发酵好的样品,去其肠衣,用捣碎机磨碎后,精确称取1 g左右样品于20 mL离心管中,加入6 mL 7%的乙酸提取1 h后于4 000 r/min离心10 min,上清液中再加入4 mL无水乙醇沉淀30 min,4 000 r/min离心10 min后,上清液经旋转蒸发浓缩至干,用1 mL蒸馏水溶解后于10 000 r/min离心10 min,上清液用于测定GABA含量。GABA含量测定参照石林娟等[21]的方法,采用Berthelot比色法测定。
1.3.4.2 感官评定
选10 位食品专业老师或学生组成感官评定小组,从产品外观、组织形态、气味和口感4 个方面对鸡肉肠进行感官评分,感官评分标准参照畅阳等[22]方法,有所改动,具体见表1。
表1 鸡肉肠感官评分标准
Table1 Criteria for sense evaluation of chicken sausage
1.4 数据统计与分析
实验设3 次重复,结果以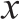 ±s来表示。采用Design-Expert 8.0.6响应面软件和 SPSS 16.0数据处理软件分析数据,进行方差分析和显著性检验。
±s来表示。采用Design-Expert 8.0.6响应面软件和 SPSS 16.0数据处理软件分析数据,进行方差分析和显著性检验。
2.1 不同乳酸菌对发酵鸡肉肠中GABA含量的影响
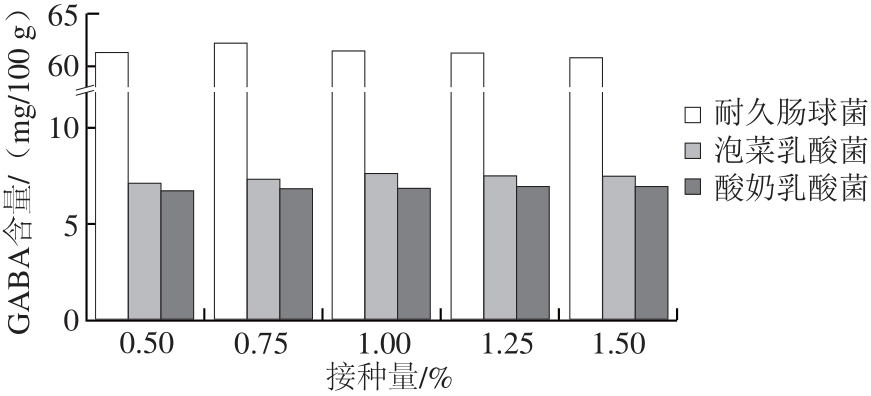
图1 不同乳酸菌对发酵鸡肉肠中GABA含量的影响
Fig.1 Effects of different lactic acid bacteria on GABA content of fermented chicken sausage
由图1可知,3 种不同乳酸菌对鸡肉肠中富集GABA的能力差异显著(P<0.05)。产GABA能力最高的菌种为耐久肠球菌,在接种量为0.75%时,GABA含量达到最高,为62.14 mg/100 g。其次是泡菜乳酸菌,在接种1%时,GABA的含量为7.6 mg/100 g;产GABA能力最差的为酸奶乳酸菌,接种1.25%时仅为6.97 mg/100 g。耐久肠球菌最高富集GABA的量是泡菜乳酸菌的8.18 倍,是酸奶乳酸菌的8.9 倍。因此,选择耐久肠球菌做为鸡肉肠富集GABA的最佳菌种。
2.2 外源添加物对发酵鸡肉肠中GABA含量和感官评分的影响
2.2.1 L-Glu对发酵鸡肉肠中GABA含量和感官评分的影响
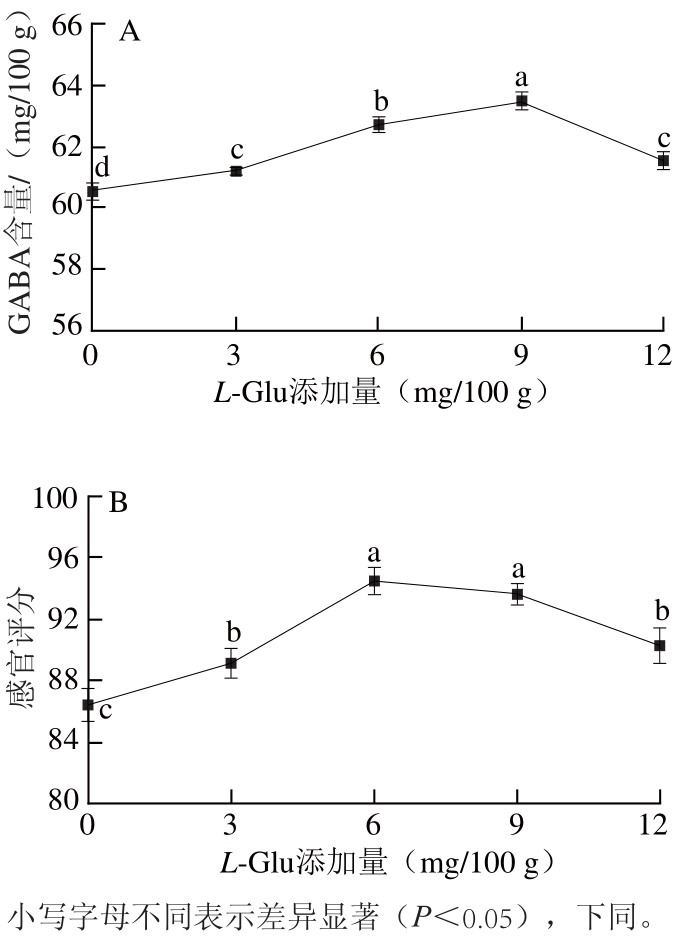
图2 2 L-Glu添加量对鸡肉肠GABA含量(A)和感官评分(B)的影响B
Fig.2 Effect of L-Glu lu addition on GABA content and sensory evaluation of chicken sausage
由图2A可见,随着L-Glu添加量的增加,发酵鸡肉肠中GABA含量呈先上升后下降的趋势。当L-Glu添加量为9 mg/100 g时,GABA含量最高,为63.47 mg/100 g,是未添加时(60.52 mg/100 g)的1.05 倍。由此说明,外源L-Glu在菌种谷氨酸脱羧酶催化下能促进GABA的合成,此结果与Bai Qingyun等[23]报道结果一致。感官评分结果表明(图2B),L-Glu添加量对产品感官有显著影响(P<0.05),随着Glu添加量的增加,感官评分呈先上升后下降的趋势,当Glu添加量在6~9 mg/100 g之间,感官评分无显著差异(P<0.05),均较高,此时,香肠口感适中,软硬合适。随后,随着Glu添加量增加,口感偏酸。综合考虑,以L-Glu添加量为9 mg/100 g为宜。
2.2.2 VB6对发酵鸡肉肠中GABA含量和感官评分的影响
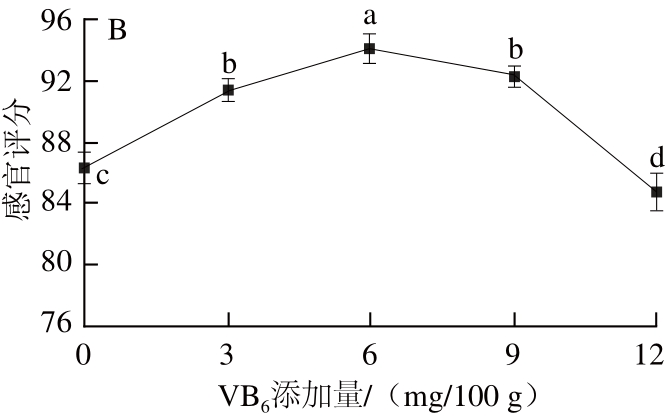
图 33 VVBB6添加量对鸡肉肠GABA含量(A)和感官评分(B)的影响
Fig.3 Effect of VB6addition on GABA content and sensory evaluation of chicken sausage
从图3A可以看出,添加VB6显著提高了发酵香肠中GABA的含量。当VB6添加量为6 mg/100 g时,GABA含量为最高,64.43 mg/100 g,是未添加组的1.07 倍。研究表明,VB6是GAD的辅基PLP的前体物质,能激活GAD,促进GABA富集[17]。随着VB6的添加,鸡肉肠感官评分先升高后降低(图3B),当添加量为6 mg/100 g时,感官评分最高,为94.1分,随后香肠颜色加深,口感也出现酸涩感,感官评分急剧下降。因此,选择VB6的添加量为6 mg/100 g。
2.2.3 CaCl2对发酵鸡肉肠中GABA含量的影响

图4 CaCl2添加量对GABA含量(A)和感官评分(B)的影响
Fig.4 Effect of CaCl2addition on GABA content and sensory evaluation of chicken sausage
如图4A所示,随着CaCl2添加量的增加,鸡肉肠中GABA含量呈先增长后下降的变化趋势。当CaCl2添加量为10 mg/100 g时,GABA富集量最大,为65.27 mg/100 g,比未添加时提高了7.8%。研究表明,无论是微生物还是植物的GAD活性受Ca2+调控,一定浓度的Ca2+促进GABA合成,过多则抑制[16,24]。感官评分表明(图4B),随着CaCl2添加量的增加,感官评分逐渐降低,这可能与Ca引起产品硬度升高、质构改变有关。CaCl2添加量不超过10 mg/100 g,感官评分均在90分以上。因此,综合考虑GABA含量和感官评分,以CaCl2添加量为10 mg/100 g为宜。
2.3 响应面试验结果
2.3.1 Box-Behnken试验设计与结果
在单因素试验基础上,采用响应面Box-Behnken设计法,建立发酵鸡肉肠GABA含量与各因素之间的数学模型,优化鸡肉肠富集GABA的最佳添加条件,试验方案和数据见表2。
表2 Box-Behnken试验设计与结果
Table2 Box-Behnken experimental design with response values

表3 回归模型方差分析
Table3 Analysis of variance (ANOVA) for the regression model
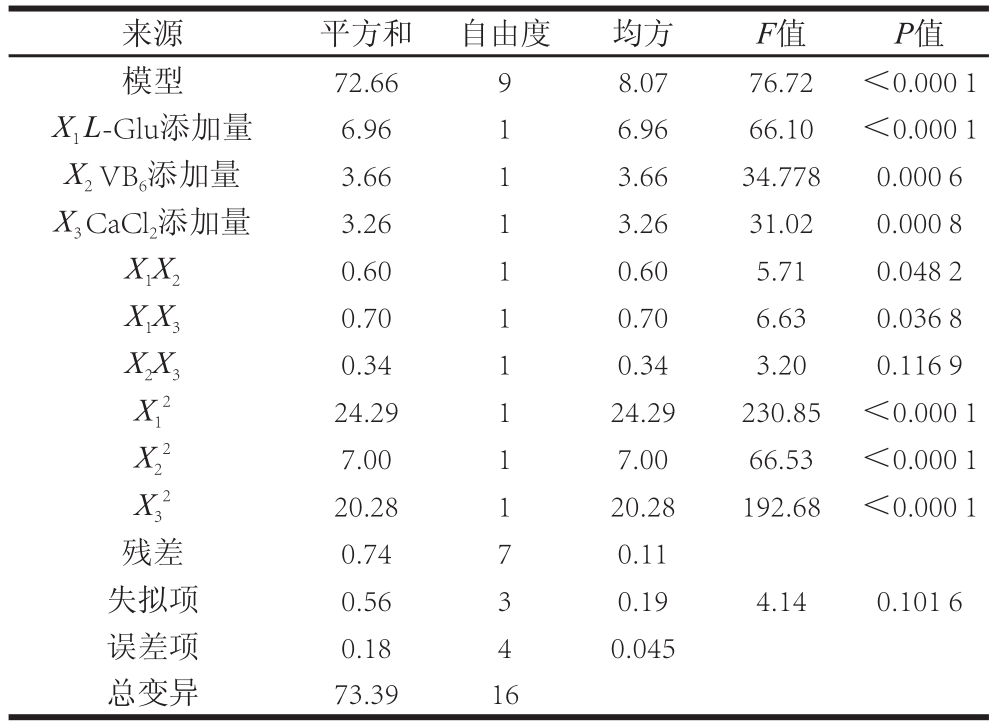
注:R2Adj=0.977 1;R2Pred=0.894 7。
用Design-Expert软件对表1中数据进行二次多元回归拟合,得到鸡肉肠中GABA含量与各变量之间的二次多项回归模型:Y=33.56+4.47X1+2.53X2+1.49X3-0.04X1X2+![]()
对回归模型进行统计分析的结果见表3,回归模型的F值为76.72,P<0.000 1,表明模型是显著的,模型的相关系数R2为0.990 0。R2越接近1,说明试验模型与真实值之间的拟合度越好。模型的信噪比为23.112(>4),表明该模型具有足够的信号强度。以上数据均证明试验模型有效[25]。
2.3.2 L-Glu、VB6和CaCl2添加量交互作用分析
根据回归法分析所得二次方程,在试验设定范围内,分析任意两变量之间的交互作用,同时固定其他变量的取值,得到二次回归方程的响应面图,见图5。
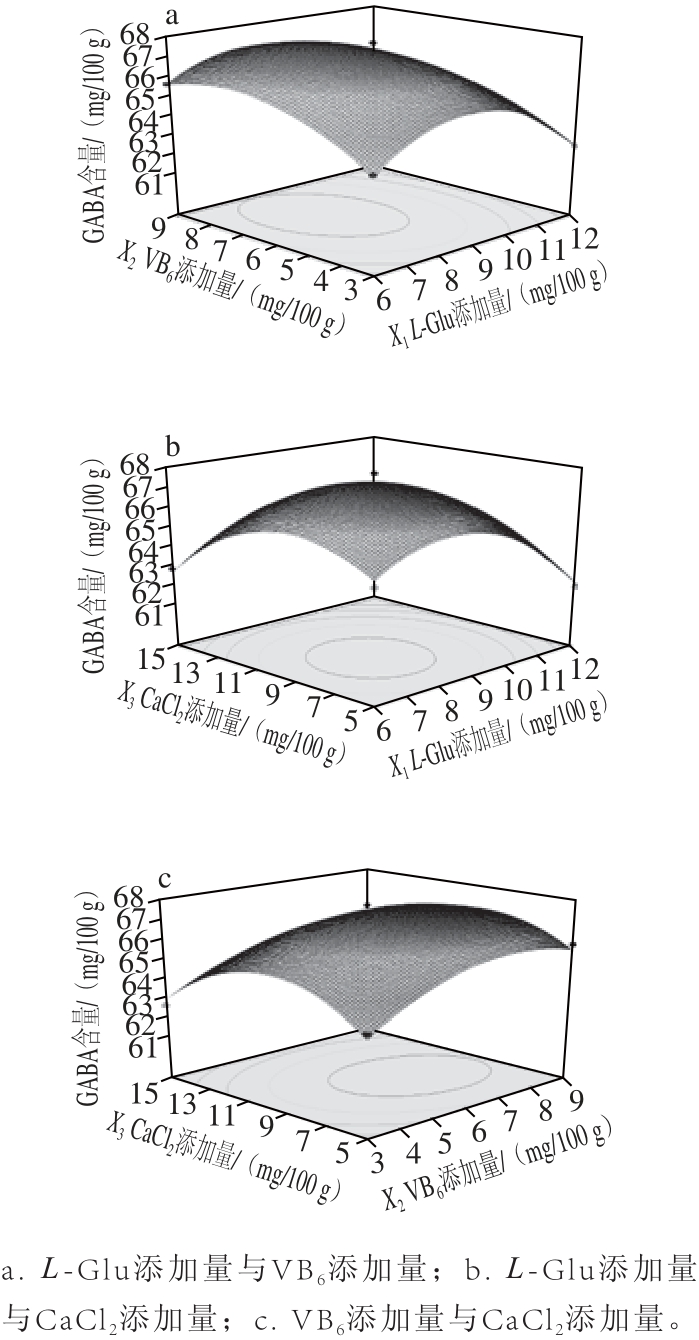
图5 两变量之间的交互作用对鸡肉肠GABA含量影响的响应面图
Fig.5 Response surface plot showing the effect of various variables on GABA content of chicken sausage
图5a表示在CaCl2添加量为10 mg/100 g条件下,L-Glu和VB6添加量对发酵鸡肉肠中GABA含量的影响。L-Glu和VB6的一次项和二次项都极显著影响发酵鸡肉肠中GABA含量,两者的交互作用影响显著(P=0.048 2)(表3)。在L-Glu添加量一定的条件下,发酵鸡肉肠中GABA含量随着VB6添加量的增加先增大后减小,在VB6添加量为6.73 mg/100 g时GABA富集最大。在VB6添加量一定的条件下,发酵鸡肉肠中GABA含量随着L-Glu添加量的增加呈先上升再下降的趋势,当L-Glu添加量达到7.75 mg/100 g时GABA含量达最大。Glu是GAD作用的底物,添加适量的Glu促进GABA合成,形成有效积累[26-27]。报道指出[28],在Lb. Paracasei NFRI 7415中GABA产量也随谷氨酸盐浓度增加而增加。
图5b表示在VB6含量为6 mg/100 g条件下,L-Glu和CaCl2添加量对发酵鸡肉肠中GABA含量的影响。CaCl2的一次项和二次项显著影响发酵鸡肉肠中GABA含量,两者的交互作用影响也显著(P=0.036 8)(表3)。L-Glu添加量一定的条件下,发酵鸡肉肠中GABA含量随着CaCl2添加量的增加先上升后下降,在添加量为8.35 mg/100 g时GABA含量最大。Ca2+通过与GAD碳末端的Ca2+/CaM调节区域结合而激活其活性[24,26],促进GABA 富集。报道指出,L-Glu与CaCl2的联合处理可促进粟谷GABA含量的提高[23],本研究结果与此一致。
图5c表示L-Glu添加量为9 mg/100 g时,VB6和CaCl2添加量对发酵鸡肉肠中GABA含量的影响,两因素的交互作用影响不显著(P=0.116 9)(表3)。在VB6添加量一定的条件下,发酵鸡肉肠中GABA含量随着CaCl2添加量的升高呈先上升后下降的趋势;当CaCl2添加量一定的条件下,发酵鸡肉肠中GABA含量随着VB6含量的增加呈先上升再下降的趋势,在添加量为6.73 mg/100 g时,发酵鸡肉肠中GABA含量达最大68.56 mg/100 g。研究表明,VB6和CaCl2联合处理促进了米糠[17]、马铃薯[29]和蚕豆[30]中GABA的积累。
2.3.3 模型优化与验证
根据响应面试验结果,乳酸菌发酵鸡肉肠富集GABA的最优外源物添加量为L-Glu 7.75 mg/100 g、VB66.73 mg/100 g、CaCl28.35 mg/100 g,鸡肉肠中GABA含量的预测值为68.56 mg/100 g。为验证预测值与真实值之间的拟合度,用优化的工艺发酵鸡肉肠富集GABA,测定其GABA含量为68.32 mg/100 g,说明实测值与预测值间非常吻合,所建模型是有效可靠的。在最优条件下鸡肉肠中GABA含量是未添加外源物中含量(62.14 mg/100 g)的1.10 倍,比普通鸡肉肠约提高10 倍。
研究不同来源乳酸菌以及添加不同外源物对发酵鸡肉肠中GABA含量的影响,并优化富集条件。结果表明,3 种乳酸菌中,耐久肠球菌发酵后GABA含量均高于泡菜乳酸菌和酸奶乳酸菌。外源添加L-Glu、VB6和CaCl2显著提高鸡肉肠中GABA含量(P<0.05),响应面优化的最佳添加量为L-Glu 7.75 mg/100 g、VB66.73 mg/100 g、CaCl28.35 mg/100 g,此时测定鸡肉肠中GABA含量为68.32 mg/100 g,与预测值吻合,最优条件下鸡肉肠中GABA含量比普通鸡肉肠约提高10 倍。方差分析表明,L-Glu、VB6、CaCl2对鸡肉肠中GABA的富集均有显著影响(P<0.05),L-Glu和VB6、L-Glu和CaCl2的交互作用均对鸡肉肠中GABA富集有显著影响(P<0.05)。
参考文献:
[1] 扶庆权, 周宏. 鸡肉火腿肠综合品质的研究[J]. 食品科学, 2011, 32(1): 45-48.
[2] 马海霞, 廖洪波, 夏扬毅, 等. 影响鸡肉发酵香肠质量的因素初探[J]. 食品与机械, 2004, 20(2): 31-33. DOI:10.13652/ j.issn.1003-5788.2004.02.018.
[3] 王恺, 慕妮, 李亮, 等. 不同发酵剂对发酵香肠挥发性风味物质的影响[J]. 食品工业科技, 2013, 34(14): 177-181. DOI:10.13386/ j.issn1002-0306.2013.14.059.
[4] 王德宝, 王佩霞, 赵丽华, 等. 复合发酵剂对羊肉发酵香肠亚硝酸盐残留量和品质的影响[J]. 食品工业科技, 2015, 36(8): 206-209. DOI:10.13386/j.issn1002-0306.2015.08.034.
[5] 李湘丽, 袁廷香, 闫吉美. 乳酸菌在发酵香肠生产过程中的应用研究进展[J]. 食品与机械, 2014, 30(6): 233-236. DOI:10.13652/ j.issn.1003-5788.2014.06.056.
[6] 白青云, 曾波, 顾振新. 低氧通气对发芽粟谷中γ-氨基丁酸含量的影响[J]. 食品科学, 2010, 31(9): 49-52.
[7] 马玉华, 王斌, 孙进, 等. γ-氨基丁酸对高脂膳食小鼠免疫功能的影响[J]. 免疫学杂志, 2014, 30(7): 599-603. DOI:10.13431/j.cnki. immunol.j.20140131.
[8] 梁金钟, 李雯, 王风青. 产γ-氨基丁酸乳酸菌的筛选及诱变育种[J]. 食品科学, 2013, 34(23): 228-232. DOI:0.7506/ spkxl002-6630-201323047.
[9] 陆小雪. 产γ-氨基丁酸乳酸菌分离与发酵条件优化及饮料开发[D].南京: 南京农业大学, 2008.
[10] 许建军. Lactococcus lactis生物合成γ-氨基丁酸及谷氨酸脱羧酶的性质研究[D]. 无锡: 江南大学, 2004.
[11] 梁金钟, 李雯, 王风青. 产γ-氨基丁酸乳酸菌的筛选及诱变育种[J].食品科学, 2013, 34(23): 228-232.
[12] SHELP B J, BOWN A W, MCLEAN M D. Metabolism and functions of gamma-aminobutyric acid[J]. Trends in Plant Science, 1999, 4: 446-452.
[13] GUO Y, CHEN H, SONG Y, et al. Effects of soaking and aeration treatment on γ-aminobutyric acid accumulation in germinated soybean (Glycine max L.)[J]. European Food Research and Technology, 2011, 232(5): 787-795. DOI:10.1007/s00217-011-1444-6.
[14] 夏江. 产γ-氨基丁酸的乳酸菌菌株筛选及其发酵条件优化[D]. 杭州:浙江大学, 2006.
[15] YANG R, YIN Y, GUO Q, et al. Purification, properties and cDNA cloning of glutamate decarboxylase in germinated faba bean (Vicia faba L.)[J]. Food Chemistry, 2013, 138(2/3): 1945-1951. DOI:10.1016/ j.foodchem.2012.11.050.
[16] 白青云. 低氧胁迫和盐胁迫下发芽粟谷γ-氨基丁酸富集机理级抗氧化性研究[D]. 南京: 南京农业大学, 2009.
[17] 张磊, 白青云, 曹晓虹, 等. 米糠富集γ-氨基丁酸的培养液组分优化[J].食品与发酵工业, 2008, 34(7): 77-81.
[18] 李蕙蕙. 鸡肉火腿肠加工过程中功能特性和蛋白质结构的初步研究[D]. 武汉: 华中农业大学, 2009.
[19] 朱建军, 王晓宇, 胡萍, 等. 组织蛋白酶对腌肉制品风味的影响[J]. 食品工程, 2013(3): 4-6. DOI:10.3969/j.issn.1673-6044.2013.03.002.
[20] 李先保, 李兴民, 南庆贤, 等. 乳酸菌发酵香肠的研制[J]. 肉类研究, 1996, 10(4): 20-24.
[21] 石林娟, 曹磊, 宋玉, 等. HPLC法和Berthelot比色法测定发芽糙米中γ-氨基丁酸的含量[J]. 粮食与饲料工业, 2015(1): 61-65. DOI:10.7633/ j/issn.1003-6202.2015.01.017.
[22] 畅阳, 王鹏, 徐幸莲, 等. 不同种木薯变性淀粉对鸡肉肠品质的影响[J]. 食品工业科技, 2012, 33(7): 58-62. DOI:10.13386/ j.issn1002-0306.2012.07.053.
[23] BAI Qingyun, CHAI Meiqing, GU Zhenxin, et al. Effects of components in culture medium on glutamate decarboxylase activity and γ-aminobutyric acid accumulation in foxtail millet (Setaria italica L.) during germination[J]. Food Chemistry, 2009, 116: 152-157. DOI:10.1016/j.foodchem.2009.02.022.
[24] BAUM G, CHEN Y, ARAZI T. A plant glutamate decarboxylase containing a calmodulin binding domain: cloning, sequence, and functional analysis[J]. Journal of Biology and Chemistry, 1993, 268:19610-19617.
[25] 朱云辉, 郭元新. 响应面法优化盐胁迫发芽苦荞富集γ-氨基丁酸的培养条件[J]. 食品科学, 2015, 36(19): 96-100. DOI:10.7506/spkx1002-6630-201519017.
[26] ZHANG H, YAO H, CHEN F, et al. Purification and characterization of glutamate decarboxylase from rice germ[J]. Food Chemistry, 2006, 101(4): 1670-1676.
[27] SSOTT-TAGGART C P, OWEN R, MICHAEL D M, et al. Regulation of γ-aminobutyric acid synthesis in situ by glutamate availability[J]. Physiologia Plantarum, 1999, 16(14): 363-369.
[28] CHUNG I, BOWN A, SHELP B. The production and efflux of 4-aminobutyrate in isolated mesophyll cells[J]. Plant Physiology, 1992, 99: 659-664.
[29] 白青云, 陈迁迁, 严煌倩, 等. 低氧通气处理马铃薯富集γ-氨基丁酸的培养条件优化[J]. 食品工业科技, 2015, 36(7): 241-245. DOI:10.13386/j.issn1002-0306.2015.07.043.
[30] 陈惠, 杨润强, 韩永斌, 等. 发芽蚕豆富集γ-氨基丁酸的培养液组分优化[J]. 中国粮油学报, 2011, 26(11): 27-31.
Optimization of Addition of Exogenous Additives for γ-Aminobutyric Acid Accumulation in Fermented Chicken Sausage
BAI Qingyun, XU Qian, LI Yuyu, LI Wen, YIN Yilin, ZHAO Li
(School of Life Science and Food Engineering, Huaiyin Institute of Technology, Huai’an 223003, China)
Abstract:The goal of this study was the accumulation of γ-aminobutyric acid (GABA) in fermented chicken sausage produced using lactic acid bacteria. Firstly, the optimal fermentation strains was selected. Then, the effects of addition of L-glutamate (L-Glu), VB6and CaCl2on the GABA content of chicken sausage were investigated, and their amounts were optimized using Box-Behnken experimental design. The results showed that among three different sources of lactic acid bacteria, the GABA-producing abilities of lactic acid bacteria from yogurt and pickles were weaker, both of which gave yields lower than10 mg/100 g, which was significantly (P < 0.05) lower than that of Enterococcus durans (62.14 mg/100 g). The addition of L-Glu, VB6and CaCl2at levels of 7.75, 6.73 and 8.35 mg/100 g, respectively was found to be optimal, yielding a GABA content of 68.32 mg/100 g, which was significantly higher than that of the samples without these additives (1.10-fold increase) and that of unfermented sausage (around 10-fold increase). Analysis of variance suggested that the regression model could quite exactly predict GABA accumulation in chicken sausage. All of the three additives had significant effects on the GABA content of chicken sausage (P < 0.01). The interaction effects between L-Glu and VB6, and between L-Glu and CaCl2were significant (P < 0.05) on GABA accumulation in chicken sausage.
Key words:fermented chicken sausage; γ-aminobutyric acid; lactic acid bacteria; optimization
DOI:10.7506/spkx1002-6630-201712032
中图分类号:TS251.5
文献标志码:A
文章编号:1002-6630(2017)12-0211-06
引文格式:白青云, 许倩, 李玉玉, 等. 发酵鸡肉肠中γ-氨基丁酸富集条件的优化[J]. 食品科学, 2017, 38(12): 211-216.
DOI:10.7506/ spkx1002-6630-201712032. http://www.spkx.net.cn
BAI Qingyun, XU Qian, LI Yuyu, et al. Optimization of addition of exogenous additives for γ-aminobutyric acid accumulation in fermented chicken sausage[J]. Food Science, 2017, 38(12): 211-216. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201712032. http://www.spkx.net.cn
收稿日期:2016-07-26
基金项目:淮安市科技计划项目(HAN2015003);淮阴工学院大学生实践创新计划项目(2016)
作者简介:白青云(1973—),女,副教授,博士,研究方向为食品科学。E-mail:baiqingyungg@163.com