
图1 变形假单胞kguK基因PCR扩增产物的琼脂糖凝胶电泳分析
Fig. 1 Agarose gel electrophoresis analysis of kguK PCR amplification products from P. plecoglossicida
何小用1,王大明2,3,孙文敬1,2,*,崔凤杰1,2,钱静亚1,2,齐向辉1,余泗莲2
(1.江苏大学食品与生物工程学院,江苏 镇江 212013;2.百勤异VC钠有限公司,江西 德兴 334221;3.江南大学生物工程学院,江苏 无锡 214122)
摘 要:采用聚合酶链式反应技术,从2-酮基葡萄糖酸工业生产菌株变形假单胞菌JUIM01中克隆2-酮基葡萄糖酸激酶的全长基因kguK,并在此基础上构建了重组菌株大肠杆菌BL21(DE3)/pET-28a(+)-kguK。在异丙基-β-D-硫代半乳糖苷的诱导下,该重组菌株表达了一个特异的分子质量约为36.0 ku融合蛋白,且该蛋白的Western-Blot鉴定结果显示为阳性。生物信息学分析结果表明,该蛋白是一种由305 个氨基酸残基组成的亲水性蛋白,定位于细胞质中,存在着与pfkB家族蛋白类似的保守结构域,其二级结构中α-螺旋、延伸链和无规则卷曲所占的比例分别为35.73%、12.79%和51.48%。
关键词:变形假单胞菌;2-酮基葡萄糖酸激酶;kguK基因;克隆;表达;生物信息学
假单胞菌(Pseudomonas)[1-4]属于氧化细菌(oxidative bacteria)[5-6],其细胞质膜的周质侧存在着与呼吸链相连的葡萄糖直接氧化系统[7-10]。该系统由膜结合的葡萄糖脱氢酶和膜结合的葡萄糖酸脱氢酶[11-14]组成,能够在细胞的周质空间中催化氧化葡萄糖为食品抗氧化剂D-异抗坏血酸合成的前体——2-酮基葡萄糖酸(2-ketogluconic acid,2KGA)[15-19]。2KGA既可积累于假单胞菌的培养液中,也可被转运至其细胞质内而被分解代谢[7,20]。因此,开展2KGA代谢机理的研究对2KGA的发酵生产具有重要的指导意义。
到目前为止,有关2KGA的分解代谢的基因调控研究主要集中于恶臭假单胞菌(P. putida)[21-22]和致病性的铜绿假单胞菌(P. aeruginosa)[23],几乎没有涉及到其他的假单胞菌,更未涉及到沙雷氏菌(Serratia)[24]、克雷伯氏菌(Klebsiella)[25-26]、醋酸杆菌(Acetobacter)[27]等能够生产2KGA的细菌菌株。相关研究结果[21-23]表明:由kguT、kguK、kguE和kguD等结构基因组成的kgu操纵子与2KGA的胞内磷酸化过程密切相关,这些基因分别编码2KGA转运蛋白(2-ketogluconate transporter,KguT)、2KGA激酶(2-ketogluconate kinase,KguK)、2KGA异构酶(2-ketogluconate epimerase,KguE)和2KGA还原酶(2-ketogluconate reductase,KguD)。另外,阻遏蛋白PtxS调控kgu操纵子的转录,同时调控其自身的合成[28-29]。
本研究以国内2KGA工业生产的主要用菌变形假单胞菌(P. plecoglossicida)JUIM01[30]为材料,对其KguK的编码基因kguK进行克隆和表达,并通过生物信息学方法分析和预测KguK蛋白的基本生物学特性,为后续的KguK蛋白纯化、鉴定及其酶学特性研究奠定基础,进而为2KGA高效生产菌株的构建提供相关的理论依据。
1.1 材料与试剂
1.1.1 菌株和质粒
变形假单胞菌(P. plecoglossicida)JUIM01 本实验室选育保藏;大肠杆菌(Escherichia coli)JM109、E. coli BL21(DE3)和质粒pET-28a(+) 本实验室保存;质粒pMD19-T 宝生物工程(大连)公司。
1.1.2 培养基
LB液体培养基:酵母提取物5.0 g/L、胰蛋白胨10.0 g/L、NaCl 10.0 g/L,pH 7.0。
LB固体培养基:在LB液体培养基中添加终质量浓度为15.0 g/L的琼脂。
1.1.3 试剂
细菌基因组提取试剂盒、2×Pfu PCR MasterMix天根生化科技(北京)有限公司;Anti-6×His antibody、HRP-conjugated Goat Anti-Rabbit lgG、W-TMB显色试剂盒、琼脂糖凝胶DNA回收试剂盒、SanPrep柱式质粒DNA小量提取试剂盒、5×Protein Loading Buffer 生工生物工程(上海)股份有限公司;限制性内切酶EcoR Ⅰ、BamH Ⅰ、T4 DNA连接酶、DNA A-Tailing Kit、DL5 000 DNA Marker、Premixed Protein Marker(Low)宝生物工程(大连)公司;PageRulerTMPrestained Protein Ladder 赛默飞世尔科技(中国)有限公司;其他试剂均为国产分析纯试剂。
1.2 仪器与设备
TGL-18M型冷冻离心机 上海卢湘仪离心机仪器有限公司;XMTD-8222型恒温水浴锅 上海精宏实验设备有限公司;C1000 TouchTM型聚合酶链式反应(polymerase chain reaction,PCR)仪、PowerPac Basic型电泳仪、GelDocTMXR+型凝胶成像仪 美国Bio-Rad公司;DYCZ-40A型电泳仪 北京市六一仪器厂;QL-901型旋涡混合器、TS-2型脱色摇床 海门市其林贝尔仪器制造有限公司;UVG20型防紫外割胶仪 上海领成生物科技有限公司;FMB40型雪花制冰机、DC-2006型低温恒温槽 上海比朗仪器有限公司;HYL-C型组合式摇床 太仓市强乐实验设备有限公司。
1.3 方法
1.3.1 变形假单胞菌JUIM01基因组DNA的提取
将低温保存的变形假单胞菌JUIM01菌株涂布于LB固体培养基上,在37 ℃恒温培养箱中培养12 h。挑取JUIM01菌株的单菌落接种于30 mL的LB液体培养基中,37 ℃、200 r/min振荡培养12 h,然后取适量菌液8 000 r/min离心收集菌体,并用无菌水洗涤菌体2 次。参照细菌基因组DNA提取试剂盒说明书,提取变形假单胞菌JUIM01基因组DNA,并在-20 ℃贮存备用。
1.3.2 PCR引物的设计与合成
根据GenBank中已公布的其他假单胞菌kguK基因的完整序列和变形假单胞菌JUIM01基因组的部分测序结果,采用同源比对的方法设计引物,设计的PCR引物如下:
P1:5’-CGGGATCCATGGCCATGTTCGTGGCCGA GCAGT-3’
P2:5’-GGAATTCTCAACCGCCGAAGCGGTCTTC GT-3’
其中,下划线部分分别为限制性内切酶BamHⅠ和EcoRⅠ的酶切位点。
引物的合成委托生工生物工程(上海)股份有限公司完成。
1.3.3 目的基因片段的扩增
以变形假单胞菌JUIM01的基因组DNA为模板,采用引物P1和P2扩增kguK基因的全长片段。
PCR体系(25.0 μL):2×Pfu PCR MasterMix 12.5 μL、引物P1(10 μmol/L)1.0 μL、引物P2(10 μmol/L)1.0 μL、基因组DNA 0.5 μL、ddH2O 10.0 μL。
PCR程序:94 ℃预变性3 min;94 ℃变性30 s,61 ℃退火30 s,72 ℃延伸1 min,反应32 个循环;72 ℃延伸5 min。1.3.4 变形假单胞菌kguK基因的克隆与测序
扩增产物经1%琼脂糖凝胶电泳分离后,参照琼脂糖凝胶DNA回收试剂盒说明书,对切胶获取的目的条带进行纯化回收。参照DNA A-Tailing Kit和pMD19-T Vector Cloning Kit说明书,将纯化得到的kguK基因片段加A尾,并进行T-A克隆,然后转化E. coli JM109的感受态细胞。在含有100 μg/mL的氨苄青霉素的LB平板(取40.0 μL的2.0 g/100 mL的X-Gal和7.0 μL的20.0 g/100mL的异丙基-β-D-硫代半乳糖苷(isopropylβ-D-thiogalactopyranoside,IPTG)混合后均匀涂布于平板表面)上进行蓝白斑筛选,挑取阳性单克隆。参照质粒DNA提取试剂盒说明书,提取重组质粒pMD19-T-kguK。对重组质粒进行双酶切(限制性内切酶BamHⅠ和EcoRⅠ)验证后,将阳性克隆子送至生工生物工程(上海)股份有限公司进行测序。
1.3.5 异源表达载体的构建与鉴定
使用限制性内切酶BamHⅠ和EcoRⅠ对重组质粒pMD19-T-kguK进行双酶切,然后采用1%琼脂糖凝胶电泳分离酶切产物;切胶获取kguK基因条带,利用琼脂糖凝胶DNA回收试剂盒回收目的DNA片段。此后,在T4 DNA连接酶作用下,将回收的目的基因片段与经过上述相同处理的表达载体pET-28a(+)连接(16 ℃连接16 h),并用该连接产物转化E. coli JM109的感受态细胞。取100 μL的转化菌液均匀涂布于含有50 μg/mL卡那霉素的LB固体培养基中,37 ℃培养12~16 h。从平板上挑取阳性单克隆,接种至含有50 μg/mL卡那霉素的LB液体培养基中,37 ℃、200 r/min振荡培养12 h。参照质粒DNA提取试剂盒说明书,提取重组质粒pET-28a(+)-kguK并进行酶切验证。
1.3.6 重组菌的构建及重组蛋白的诱导表达
利用重组质粒pET-28a(+)-kguK转化E. coli BL21(DE3)的感受态细胞,然后将转化菌液均匀涂布于含有50 μg/mL卡那霉素的LB固体培养基中,37 ℃培养12~16 h。从平板上挑取阳性单克隆至含有50 μg/mL卡那霉素的LB液体培养基中过夜振荡培养(37 ℃,200 r/min),然后以2%的接种量转接至相同的液体培养基中进行扩大培养(37 ℃,200 r/min)。当培养液的OD600nm达到0.6左右时,加入IPTG至终浓度为0.8 mmol/L,在20 ℃、200 r/min的条件下诱导表达12 h。
1.3.7 SDS-PAGE分析与Western-Blot鉴定
取经过诱导的细胞培养液1.0 mL,8 000 r/min离心2 min收集菌体,并用预冷的无菌PBS缓冲液洗涤2 次,然后使菌体重悬于100 μL的PBS缓冲液中。取20.0 μL的重悬菌液与5.0 μL的十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)蛋白上样缓冲液充分混匀,于100 ℃水中煮沸5 min。待样品冷却后,对其全细胞蛋白进行SDS-PAGE分析。
以Anti-6×His antibody(抗6×His单克隆兔抗)为一抗、HRP-conjugated Goat Anti-Rabbit lgG(IgG/HRP羊抗兔)为二抗,以预染蛋白质分子质量标准PageRulerTMPrestained Protein Ladder为Marker,对经SDS-PAGE分离的KguK融合蛋白进行Western-Blot鉴定。
1.3.8 变形假单胞菌KguK蛋白的序列分析
采用DNAMAN软件进行基因序列和氨基酸序列的分析比对;使用ExPASy的ProtParam工具对kguK编码的蛋白质KguK的理化性质进行预测和分析;应用ExPASy的ProtScale工具在线分析蛋白的疏水性;应用TMHMM Server v. 2.0、SignalP 4.1 Server、Kinasephos和PSORT WWW Server中的PSORT Prediction在线工具分别预测目的基因编码蛋白的跨膜结构域、信号肽、激酶磷酸化修饰位点和亚细胞定位;利用PSIPRED V3.3在线分析工具分析蛋白的二级结构;采用ExPASy中COILS工具预测KguK蛋白的卷曲螺旋结构;使用InterProScan在线工具对KguK蛋白的保守结构域和功能位点进行分析;利用MEGA6.0软件构建蛋白的系统进化树;应用Swiss-Model在线工具进行同源模建,并用PDBsum Generate在线蛋白检测工具对预测的蛋白模型进行评估。
2.1 变形假单胞菌kguK基因片段的克隆
图1 变形假单胞kguK基因PCR扩增产物的琼脂糖凝胶电泳分析
Fig. 1 Agarose gel electrophoresis analysis of kguK PCR amplification products from P. plecoglossicida
以变形假单胞菌JUIM01的基因组为模板,以P1和P2为引物进行PCR扩增,得到了一段长度约为900 bp的核酸片段(图1)。
采用DNA A-Tailing Kit对切胶回收的PCR产物进行处理后,经连接、转化、蓝白斑筛选和质粒提取等步骤,得到了重组质粒pMD19-T-kguK。在该质粒双酶切产物的琼脂糖凝胶电泳图谱(图2)中,存在2 条得到明显分离的条带,其中一条在大约900 bp处,另一条在大约2 700 bp处,与预期的结果一致。上述结果表明,重组质粒pMD19-T-kguK得到了正确的构建。

图2 重组质粒pMD19-T-kguK双酶切产物的琼脂糖凝胶电泳分析
Fig. 2 Electrophoretic analysis of the restriction enzyme-digested products of recombinant plasmid pMD19-T-kguK

图3 变形假单胞KguK的基因序列及氨基酸序列
Fig. 3 Gene and amino acid sequences of KguK from P. plecoglossicida
由图3可知,克隆的基因片段的核苷酸序列长度为918 bp。DNAMAN分析结果表明,克隆的基因片段具有一个完整开放阅读框,其起始密码子为ATG,终止密码子为TGA。利用BLAST进行同源性比对发现,本研究克隆的基因片段与P. putida NBRC 14164、P. putida KT2440、P. aeruginosa UCBPP-PA14、P. aeruginosa NCGM 1900和P. fluorescens F113的编码KguK的基因片段在核苷酸序列上的一致性分别为71.91%、71.59%、67.61%、67.51%和64.37%,初步认为该序列为变形假单胞菌JUIM01编码KguK的基因序列。本研究克隆的变形假单胞菌JUIM01的kguK基因序列已被递交至GenBank数据库,登录号为KU168043。
2.2 异源表达载体的构建
对重组质粒pMD19-T-kguK和表达载体pET-28a(+)分别进行双酶切处理,经电泳分离、DNA回收、连接、转化、抗性筛选和质粒提取等步骤,得到了重组质粒pET-28a(+)-kguK。1%琼脂糖凝胶电泳分析结果(图4)表明:重组质粒pET-28a(+)-kguK的单酶切(BamHⅠ处理)产物显示为长度约为6 300 bp的单一条带,与预期的结果相符;重组质粒pET-28a(+)-kguK的双酶切(BamHⅠ和EcoRⅠ处理)产物显示为长度分别为5 400 bp左右和900 bp左右的2 条条带,与预期的结果一致。上述研究结果证明,基因kguK已成功构建到表达载体pET-28a(+)-kguK中。

图4 重组质粒pET-28a(+)-kguK酶切产物的琼脂糖凝胶电泳分析
Fig. 4 Electrophoretic analysis of the restriction enzyme-digested products of recombinant plasmid pET-28a(+)-kguK
2.3 重组菌的构建及重组蛋白的诱导表达

图5 重组菌株E. coli BL21(DE3)/pET-28a(+)-kguK全细胞蛋白的SDS-PAGE分析
Fig. 5 SDS-PAGE analysis of the whole-cell proteins extracted from E. coli BL21(DE3)/ pET-28a(+)-kguK
利用重组质粒pET-28a(+)-kguK转化E. coli BL21(DE3)的感受态细胞,经抗性筛选得到了重组菌E. coli BL21(DE3)/pET-28a(+)-kguK。SDS-PAGE分析结果(图5)表明:在相同的诱导条件下,重组菌E. coli BL21(DE3)/pET-28a(+)-kguK表达出分子质量约为36.0 ku的融合蛋白(其中标签蛋白的分子质量约为3.5 ku),而E. coli BL21(DE3)和E. coli BL21(DE3)/ pET-28a(+)的全细胞蛋白中没有该蛋白的存在。
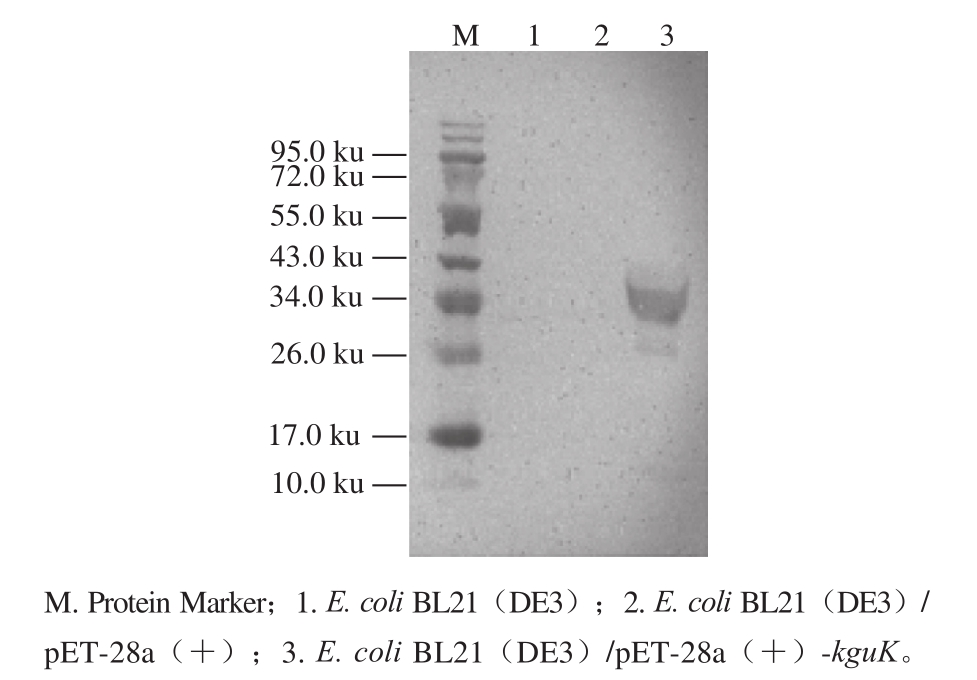
图6 重组菌株E. coli BL21(DE3)/pET-28a(+)-kguK表达的融合蛋白的Western-Blot分析
Fig. 6 Western-Blot analysis of the protein expressed by E. coli BL21(DE3)/pET-28a(+)-kguK
对经过SDS-PAGE分离的重组蛋白进行Western-Blot鉴定,结果如图6所示。E. coli BL21(DE3)/pET-28a(+)-kguK表达的融合蛋白的鉴定结果显示为阳性,进一步证明了变形假单胞菌JUIM01的kguK基因在E. coli BL21(DE3)中的成功表达。
2.4 变形假单胞菌KguK蛋白的生物信息学分析
2.4.1 变形假单胞菌KguK的基本理化性质预测
采用ExPASy的ProtParam工具进行的分析结果表明:KguK蛋白由305 个氨基酸残基组成,其中带负电荷的氨基酸残基(Asp+Glu)总数为37 个,带正电荷的氨基酸残基(Arg+Lys)为31 个。该蛋白的理论分子质量和等电点分别为32.5 ku和5.47,分子式为C1417H2265N423O429S14。KguK蛋白在溶液中的不稳定指数为40.78(>40),说明该蛋白在溶液中性质不稳定。KguK的脂溶性指数为92.20,总平均亲水指数为-0.032,预测该蛋白为亲水性蛋白,但亲水性较低。采用ExPASy的ProtScale工具对KguK蛋白进行疏水性分析发现,该蛋白为亲水性蛋白,预测结果与上述结果一致。
2.4.2 变形假单胞菌KguK的跨膜结构、细胞定位、信号肽及潜在激酶磷酸化位点预测
采用TMHMM Server v. 2.0进行的预测显示,KguK蛋白无跨膜结构域,为非跨膜蛋白。利用PSORT Prediction进行的预测结果表明,KguK蛋白定位于细胞质中。采用SignalP 4.1 Server对KguK蛋白的氨基酸序列进行N端信号肽的预测结果显示,该蛋白无信号肽。
采用NetPhos2.0在线分析工具对KguK蛋白中潜在的苏氨酸、丝氨酸和酪氨酸的激酶磷酸化位点进行分析,结果显示:KguK蛋白中可能存在10 个丝氨酸激酶磷酸化修饰位点(第50、99、100、103、106、130、132、150、160、283位氨基酸)、4 个苏氨酸激酶磷酸化修饰位点(第167、193、197、230位氨基酸)和2 个酪氨酸激酶磷酸化修饰位点(第223、299位氨基酸)。
2.4.3 变形假单胞菌KguK的二级结构及卷曲螺旋预测
通过PSIPRED V3.3在线分析工具对KguK蛋白的二级结构进行了预测,结果显示:KguK蛋白二级结构中具有α-螺旋、延伸链以及无规则卷曲3 种结构,所占比例分别为35.73%、12.79%和51.48%。
卷曲螺旋是蛋白质中2~7 条α-螺旋链互相缠绕形成类似麻花状结构的总称,存在于转录因子、膜蛋白、酶等蛋白质中,具有分子识别、膜通道和代谢调控等生物学功能。使用COILS server预测了蛋白KguK的卷曲螺旋结构,结果显示:在3 种不同的窗口宽度下的检测值均较低,表明KguK蛋白中可能不存在卷曲螺旋结构。
2.4.4 变形假单胞菌KguK的结构域和功能位点分析
InterPro数据库整合了蛋白家族、结构域和功能位点等数据库资源,如SMART、Pfam、TIGRFAMs、ProDom、PROSITE、PRINTS等常用数据库,检测结果的准确度较高。使用InterProScan在线工具分析KguK蛋白的结构域和功能位点,结果表明,KguK蛋白存在PfkB蛋白结构域,具有磷酸转移酶活性,受体基团为醇基,其保守位点位于第244~257个氨基酸残基之间。
2.4.5 变形假单胞菌KguK的系统进化分析

图7 KguK蛋白的系统进化树
Fig. 7 Phylogenetic tree of KguK proteins from Pseudomonas
通过BLAST搜索,从NCBI数据库中获得了变形假单胞菌KguK的同源序列,然后利用ClustW软件进行多序列同源性多重比对,采用MEGA6.0软件对7 个不同来源的氨基酸序列进行系统进化分析。分析结果(图7)表明,变形假单胞菌JUIM01的KguK与P. putida NBRC 14164的KguK的同源性较高,在氨基酸序列上的一致性达73.82%。
2.4.6 变形假单胞菌KguK的三级结构预测与检验
利用Swiss-Model在线预测工具,采用自动模式对变形假单胞菌JUIM01的KguK蛋白的三级结构进行同源模建,选取与KguK蛋白同源性最高的PfkB蛋白(4du5.1A)为模板,模拟KguK蛋白的三级结构,结果如图8所示。
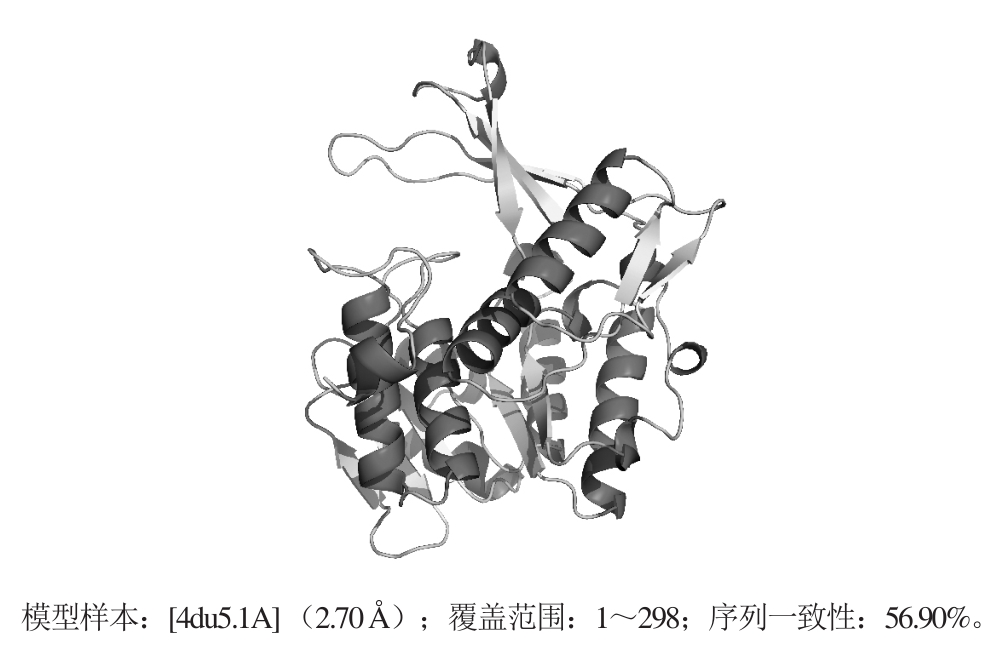
图8 变形假单胞菌JUIM01蛋白KguK的三级结构
Fig. 8 Tertiary structure of KguK from P. plecoglossicida JUIM01 predicted by Swiss-model

图9 变形假单胞菌JUIM01蛋白KguK的拉氏构象图
Fig. 9 Ramachandran diagram of KguK from P. plecoglossicida JUIM01
采用PDBsum Generate工具对预测结果进行评估,结果如图9所示。根据构象的稳定性,拉氏构象图被分为4 个区域,即最佳区、次允许区、一般区和不允许区。一般认为,最佳区达90%以上、不允许区低于1%的模型结构的稳定性较好。对变形假单胞菌KguK三级结构预测模型的评估结果显示,上述4 个区域的统计数据分别为93.7%、6.3%、0.0%和0.0%,表明所构建的KguK蛋白三级结构模型的质量较高。
采用PCR技术,从工业生产菌株变形假单胞菌JUIM01克隆了与2KGA代谢密切相关的KguK的全长基因,其核苷酸序列长度为918 bp。利用所克隆的基因kguK,构建了重组菌E. coli BL21(DE3)/pET-28a(+)-kguK。在IPTG的诱导下,该重组菌表达了一个特异性的分子质量约为36.0 ku的融合蛋白(其中标签蛋白的分子质量约为3.5 ku)。该重组蛋白的Western-Blot鉴定结果显示为阳性,进一步证明了变形假单胞菌JUIM01的kguK基因在E. coli BL21(DE3)中的成功表达。
生物信息学分析结果表明:变形假单胞菌KguK是由305 个氨基酸残基组成的亲水性蛋白,理论分子质量为32.5 ku,定位于细胞质中,无跨膜结构和信号肽。变形假单胞菌KguK存在一个保守结构域,与pfkB家族蛋白的保守结构域类似。在该蛋白的氨基酸序列上,存在着16 个潜在的激酶磷酸化修饰位点。在该蛋白的二级结构中,α-螺旋、延伸链和无规则卷曲3 种结构形式所占的比例分别为35.73%、12.79%和51.48%。
参考文献:
[1] PALLERONI N J. The Pseudomonas story[J]. Environmental Microbiology, 2010, 12(6): 1377-1383. DOI:10.1111/j.1462-2920.2009.02041.x.
[2] LA ROSA R, NOGALES J, ROJO F. The Crc/CrcZ-CrcY global regulatory system helps the integration of gluconeogenic and glycolytic metabolism in Pseudomonas putida[J]. Environmental Microbiology, 2015, 17(9): 3362-3378. DOI:10.1111/1462-2920.12812.
[3] KALRA S, MIDHA K, KAUR A. Production, purification and characterization of L-asparaginase from Pseudomonas aeruginosa 2488 using fermentation[J]. International Journal of Advanced Research, 2016, 4(1): 541-549.
[4] SUN Wenjing, XIAO Fangfang, WEI Zhuan, et al. Non-sterile and buffer-free bioconversion of glucose to 2-keto-gluconic acid by using Pseudomonas fluorescens AR4 free resting cells[J]. Process Biochemistry, 2015, 50(4): 493-499. DOI:10.1016/ j.procbio.2015.01.011.
[5] MATSUSHITA K, OHNO Y, SHINAGAWA E, et al. Membranebound, electron transport-linked, D-glucose dehydrogenase of Pseudomonas fluorescens. Interaction of the purified enzyme with ubiquinone or phospholipid[J]. Agricultural and Biological Chemistry, 1982, 46(4): 1007-1011. DOI:10.1080/00021369.1982.10865179.
[6] GILARDI G L, FAUR y C. Pseudomonas mesophilica and an unnamed taxon, clinical isolates of pink-pigmented oxidative bacteria[J]. Journal of Clinical Microbiology, 1984, 20(4): 626-629.
[7] DEL CASTILLO T, RAMOS J L, RODRIGUEZ-HERVA J J, et al. Convergent peripheral pathways catalyze initial glucose catabolism in Pseudomonas putida: genomic and flux analysis[J]. Journal of Bacteriology, 2007, 189(14): 5142-5152. DOI:10.1128/JB.00203-07.
[8] DADDAOUA A, MOLINA-SANTIAGO C, DE LA TORRE J, et al. GtrS and GltR form a two-component system: the central role of 2-ketogluconate in the expression of exotoxin A and glucose catabolic enzymes in Pseudomonas aeruginosa[J]. Nucleic Acids Research, 2014, 42(12): 7654-7663. DOI:10.1093/nar/gku496.
[9] SHRIVASTAVA R, BASU B, GODBOLE A, et al. Repression of the glucose-inducible outer-membrane protein OprB during utilization of aromatic compounds and organic acids in Pseudomonas putida CSV86[J]. Microbiology, 2011, 157(5): 1531-1540. DOI:10.1099/ mic.0.047191-0.
[10] PANDEy S, MODAK A, PHALE P S, et al. Cloning, purification, crystallization and preliminary X-ray diffraction studies of periplasmic glucose binding protein of Pseudomonas putida CSV86[J]. Advances in Bioscience and Biotechnology, 2015, 6(5): 164-171. DOI:10.4236/ abb.2015.63016.
[11] MATSUSHITA K, OHNO y, SHINAGAWA E, et al. Membrane-bound D-glucose dehydrogenase from Pseudomonas sp.: solubilization, purification and characterization[J]. Agricultural and Biological Chemistry, 1980, 44(7): 1505-1512. DOI:10.1271/bbb1961.44.1505.
[12] MATSUSHITA K, AMEyAMA M. D-Glucose dehydrogenase from Pseudomonas fluorescens, membrane-bound[J]. Methods in Enzymolog, 1982, 89(4): 149-154. DOI:10.1016/S0076-6879(82)89026-5.
[13] KUMAR C, yADAV K, ARCHANA G, et al. 2-Ketogluconic acid secretion by incorporation of Pseudomonas putida KT2440 gluconate dehydrogenase (gad) operon in Enterobacter asburiae PSI3 improves mineral phosphate solubilization[J]. Current Microbiology, 2013, 67(3): 388-394. DOI:10.1007/s00284-013-0372-z.
[14] MATSUSHITA K, SHINAGAWA E, ADACHI O, et al. Membranebound D-gluconate dehydrogenase from Pseudomonas aeruginosa: purification and structure of cytochrome-binding form[J]. Journal of Biochemistry, 1979, 85(5): 1173-1181. DOI:10.1093/oxfordjournals. jbchem.a132441.
[15] 周强, 魏转, 孙文敬, 等. D-异抗坏血酸生产技术研究进展[J]. 食品科学, 2008, 29(8): 647-651. DOI:10.3321/j.issn:1002-6630.2008.08.158.
[16] PAPPENBERGER G, HOHMANN H P. Industrial production of L-ascorbic acid (vitamin C) and D-isoascorbic acid[J]. Advances in Biochemical Engineering Biotechnology, 2014, 143: 143-188. DOI:10.1007/10_2013_243.
[17] 张抗, 史小利, 龚舒, 等. D-异抗坏血酸钠合成工艺研究[J]. 低碳世界, 2014(10): 351-352.
[18] 魏转, 余泗莲, 孙文敬, 等. 2-酮基-D-葡萄糖酸发酵生产研究进展[J]. 食品科学, 2008, 29(8): 636-639. DOI:10.3321/ j.issn:1002-6630.2008.08.155.
[19] UMEZAWA K, TAKEDA K, ISHIDA T, et al. A novel pyrroloquinoline quinone-dependent 2-keto-D-glucose dehydrogenase from Pseudomonas aureofaciens[J]. Journal of Bacteriology, 2015, 197(8): 1322-1329. DOI:10.1128/JB.02376-14.
[20] SUN Wenjing, ZHOU yanzheng, ZHOU Qiang, et al. Semi-continuous production of 2-keto-gluconic acid by Pseudomonas fluorescens AR4 from rice starch hydrolysate[J]. Bioresource Technology, 2012, 110: 546-551. DOI:10.1016/j.biortech.2012.01.040.
[21] NELSON K E, WEINEL C, PAULSEN I T, et al. Complete genome sequence and comparative analysis of the metabolically versatile Pseudomonas putida KT2440[J]. Environmental Microbiology, 2002, 4(12): 799-808. DOI:10.1046/j.1462-2920.2002.00366.x.
[22] NIKEL P I, CHAVARRIA M, FUHRER T, et al. Pseudomonas putida KT2440 metabolizes glucose through a cycle formed by enzymes of the entner-doudoroff, embden-meyerhof-parnas, and pentose phosphate pathways[J]. Journal of Biological Chemistry, 2015, 290(43): 25920-25932. DOI:10.1074/jbc.M115.687749.
[23] SWANSON B L, HAGER P, PHIBBS P, et al. Characterization of the 2-ketogluconate utilization operon in Pseudomonas aeruginosa PAO1[J]. Molecular Microbiology, 2000, 37(3): 561-573. DOI:10.1046/j.1365-2958.2000.02012.x.
[24] MISENHEIMER T J, ANDERSON R F, LAGODA A A, et al. Production of 2-ketogluconic acid by Serratia marcescens[J]. Applied Microbiology, 1965, 13(3): 393-396.
[25] WEI Dong, XU Jiqing, SUN Junson, et al. 2-Ketogluconic acid production by Klebsiella pneumoniae CGMCC 1.6366[J]. Journal of Industrial Microbiology and Biotechnology, 2013, 40(6): 561-570. DOI:10.1007/s10295-013-1261-y.
[26] SUN yuehong, WEI Dong, SHI Jiping, et al. Two-stage fermentation for 2-ketogluconic acid production by Klebsiella pneumoniae[J]. Journal of Microbiology and Biotechnology, 2014, 24(6): 781-787. DOI:10.4014/jmb.1401.01038.
[27] ŠVITEL J, ŠTURDIK E. 2-Ketogluconic acid production by Acetobacter pasteurianus[J]. Applied Biochemistry and Biotechnology, 1995, 53(1): 53-63. DOI:10.1007/BF02783481.
[28] DADDAOUA A, KRELL T, ALFONSO C, et al. Compartmentalized glucose metabolism in Pseudomonas putida is controlled by the PtxS repressor[J]. Journal of Bacteriology, 2010, 192(17): 4357-4366. DOI:10.1128/JB.00520-10.
[29] DADDAOUA A, FILLET S, FERNANANDEZ M, et al. Genes for carbon metabolism and the toxA virulence factor in Pseudomonas aeruginosa are regulated through molecular interactions of PtxR and PtxS[J]. PLoS ONE, 2012, 7(7): e39390. DOI:10.1371/journal. pone.0039390.
[30] 孙文敬, 栾方, 王大明, 等. 变形假单胞菌2-酮基葡萄糖酸转运蛋白基因的克隆与分析[J]. 现代食品科技, 2016, 32(6): 50-55. DOI:10.13982/j.mfst.1673-9078.2016.6.009.
Cloning, Expression and Bioinformatic Analysis of 2-Ketogluconate Kinase Gene from Pseudomonas plecoglossicida
HE Xiaoyong1, WANG Daming2,3, SUN Wenjing1,2,*, CUI Fengjie1,2, QIAN Jingya1,2, QI Xianghui1, YU Silian2
(1. School of Food and Biological Engineering, Jiangsu University, Zhenjiang 212013, China; 2. Parchn Sodium Isovitamin C Co. Ltd., Dexing 334221, China; 3. School of Biotechnology, Jiangnan University, Wuxi 214122, China)
Abstract:The full-length sequence of 2-ketegluconate kinase gene (kguK) from was cloned from an industrial 2KGA producer of Pseudomonas plecoglossicida JUIM01 and expressed in the recombinant strain Escherichia coli BL21(DE3)/ pET-28a(+)-kguK with isopropyl β-D-1-thiogalactopyranoside induction. The specific fusion protein KguK had a molecular weight of about 36.0 ku, which was confirmed by Western-Blot. The bioinformatics analysis showed that KguK in P. plecoglossicida was a hydrophilic protein with 305 amino acid residues. It was located in the cytoplasm, having a conserved domain similar to that of the pfkB family. The predicted secondary structure contained 35.73% of α-helixes, 12.79% of extended strand and 51.48% of random coil.
Key words:Pseudomonas plecoglossicida; 2-ketegluconate kinase (KguK); kguK gene; cloning; expression; bioinformatics
DOI:10.7506/spkx1002-6630-201716002
中图分类号:Q781
文献标志码:A
文章编号:1002-6630(2017)16-0010-07
引文格式:
何小用, 王大明, 孙文敬, 等. 变形假单胞菌2-酮基葡萄糖酸激酶基因的克隆、表达及生物信息学分析[J]. 食品科学, 2017, 38(16): 10-16. DOI:10.7506/spkx1002-6630-201716002. http://www.spkx.net.cn
HE Xiaoyong, WANG Daming, SUN Wenjing, et al. Cloning, expression and bioinformatic analysis of 2-ketogluconate kinase gene from Pseudomonas plecoglossicida[J]. Food Science, 2017, 38(16): 10-16. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201716002. http://www.spkx.net.cn
收稿日期:2016-11-08
基金项目:国家自然科学基金面上项目(31571885);国家高技术研究发展计划(863计划)项目(2012AA022103);江西省科技计划项目(赣知发[2015]64号);江西省创新团队建设计划项目(20142BCB24024);江西省科技平台建设计划项目(2010DTZ01900);德兴市科技计划项目(德科发[2015]44号)
作者简介:何小用(1989—),男,硕士研究生,研究方向为食品生物技术。E-mail:hxy9808@126.com
*通信作者:孙文敬(1964—),男,研究员,博士,研究方向为工业生物技术。E-mail:juswj@163.com