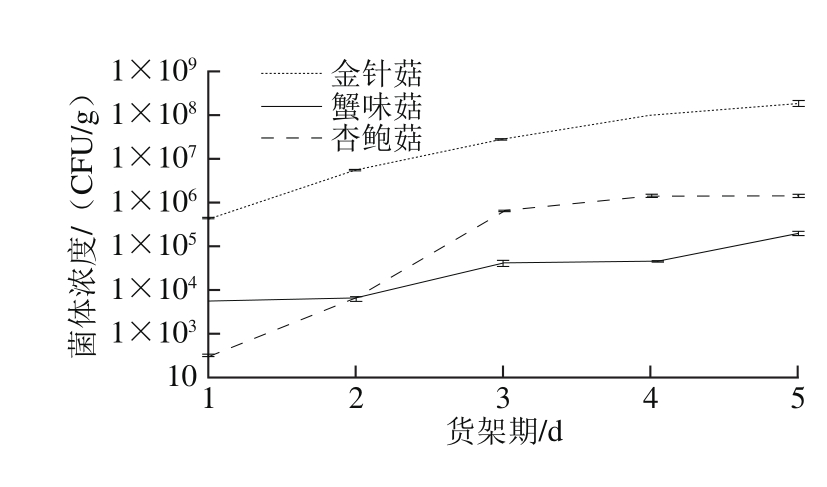
图1 3 种食用菌货架期外源细菌数量变化
Fig. 1 Changes in total number of exogenous bacteria in three kinds of mushrooms during storage
王琼英,高 巍,赵梦然,邬向丽,陈 强,曲积彬,黄晨阳*
(中国农业科学院农业资源与农业区划研究所,农业部农业微生物资源收集与保藏重点实验室,北京 100081)
摘 要:目前,市售食用菌鲜品大多保藏于常温条件下,食用菌因营养丰富、表面组织鲜嫩而更易滋生外源微生物。以市售常温保藏的金针菇、杏鲍菇、蟹味菇鲜品为材料,分析货架期内其外源细菌数量、种类、群落多样性的变化。结果表明,3 种食用菌在货架期内外源细菌数量明显增加,货架期5 d蟹味菇增长了29 倍,金针菇、杏鲍菇分别增长了417、4 116 倍。外源细菌种类及相对丰度在货架期呈动态变化,表现为优势菌在货架期1~5 d能够被检测到,蟹味菇假单胞菌属丰度增加和金针菇乳球菌属丰度降低。Shannon指数表明在货架期内,金针菇外源细菌多样性明显低于杏鲍菇和蟹味菇。主成分分析结果表明,3 种食用菌各自在不同货架期内菌群结构呈现相似性,蟹味菇和杏鲍菇外源细菌菌群结构存在一定的相似性,金针菇外源细菌菌群结构与蟹味菇和杏鲍菇存在差异。
关键词:货架期;高通量测序法;多样性;致病菌;腐败菌
食用菌兼具营养、美味和保健功能,深受消费者青睐。据中国食用菌协会统计,2015年我国食用菌总产量3 476.15万 t,产值2 516.38亿 元。市售食用菌以干品和鲜品为主,随着消费需求的改变,食用菌鲜品占比日益扩大[1]。食用菌生产环节易于控制,严把原料质量、规范栽培管理即可完全避免重金属污染、农药残留等问题[2],但是市售食用菌鲜品存在质量的问题,主要原因是采后的包装、贮运、销售环节采用常温,未能有效防范外源细菌的繁殖[3-5]。研究食用菌外源细菌数量、种类、群落多样性的变化成为必然。
国外学者对食用菌外源细菌开展初步研究。Reinartz等[6]对-24 ℃冻存一年的双孢蘑菇切片和其他食品进行微生物区系检测,以探究微生物死亡率以及不同细菌的生存能力,发现肠杆菌科细菌显著减少,乳酸细菌、乳酸杆菌、假单胞菌、酵母菌和霉菌也有相当程度的减少。Leff等[7]对双孢蘑菇和蔬菜、水果等进行外源细菌研究,发现双孢蘑菇外源细菌中肠杆菌科细菌比例很小,其外源细菌主要隶属于根瘤菌科、微球菌科、鞘脂杆菌科、丛毛单胞菌科,且传统农业式栽培比有机农业式栽培外源细菌丰度更高。Jaworska等[8]对新鲜未贮存、20 ℃贮存48 h、4 ℃贮存48 h和4 ℃贮存96 h的牛肝菌样品进行外源微生物分析,结果发现,20 ℃贮存48 h的样品可以检测到大量乳酸菌,还包括一些芽孢菌、霉菌和酵母,已不适合销售;即使经过热烫处理后,并没有明显减少上述微生物数量;但4 ℃贮存96 h的牛肝菌未检测到。国内对食用菌外源细菌的研究鲜有报道。
本研究以市售金针菇、杏鲍菇、蟹味菇鲜品为对象,分析常温条件下外源细菌数量、种类、群落多样性,探寻货架期内外源细菌动态变化规律,以期为保障食用菌鲜品质量安全提供科学依据。
1.1 材料与试剂
金针菇、杏鲍菇、蟹味菇3 种食用菌为材料,购自北京市本地超市,常温销售,当天上架,托盘独立包装。
LB培养基 生工生物工程(上海)股份有限公司;细菌基因组提取试剂盒 天根生化科技(北京)有限公司。
1.2 仪器与设备
Shock-Mixer1脉冲均质器 广东环凯微生物科技有限公司;C1000聚合酶链式反应(polymerase chain reaction,PCR)仪 美国Bio-Rad公司。
1.3 方法
1.3.1 货架期设置
金针菇、杏鲍菇、蟹味菇3 种食用菌各购买25 份,选择当天上架的、独立包装的同批鲜品,置于常温条件下存放。货架期设置1~5 d共5 个处理,当天随机取样3 份记为货架期1 d。每次随机取样,3 个重复。
1.3.2 样品制备
处理过程中采样原则、采样方案参照GB 4789.1—2016《食品微生物学检验 总则》[9]。采样后,样品预处理:经无菌操作将新鲜子实体切碎,混合均匀,取25.0 g放入无菌袋中,加入225.0 mL质量分数0.85% NaCl溶液,经脉冲匀质器振荡30 s,均质液即为10-1样品稀释液,以质量分数0.85% NaCl溶液依次制备10-2、10-3、10-4、10-5、10-6梯度稀释液,备用。
1.3.3 菌种鉴定
分离、纯化:选择10-2、10-3、10-4、10-5、10-6多个梯度样品稀释液涂平板,选择适宜浓度的平板,根据平板上菌落生长的形态、大小、颜色等特征挑取单菌落,进行划线培养。
菌落计数:计数原则、方案参照GB 4789.2—2016《食品微生物学检验 菌落总数测定》[10]。
鉴定:用煮沸法提取细菌DNA(取菌苔于50 µL ddH2O中,100 ℃煮沸15 min,-20 ℃放置90 s,12 000 r/min离心2 min,上清液备用)。使用通用引物27F(5’-AGAGTTTGATCCTGGCTCAG-3’)和1492R(5’-TACGGTTACCTTGTTACGACTT-3’)扩增16S rDNA全长[11]。扩增体系为:2 µL DNA模板,5 µL 10×ExTaq Buffer,2 µL dNTPs,引物27F和1492R各0.75 µL(10 µmol/L),0.25 µL ExTaq聚合酶,ddH2O补足至25 µL。PCR产物电泳检测后,送北京博迈德公司测序,测序结果与Eztaxon数据库比对得到相似度最高的种[12]。
1.3.4 16S rDNA高通量测序
取200 mL洗脱液,分装于50 mL离心管,12 000 r/min离心20 min,弃上清液,用1.0 mL无菌水悬浮沉淀,合并于10 mL离心管。然后,12 000 r/min离心1 min,弃上清液,沉淀用于DNA提取。
基因组提取:使用细菌基因组提取试剂盒,依据说明书操作,所提DNA保存于-20 ℃。
16S rDNA高通量测序:委托北京奥维森公司进行,以338F(5’-ACTCCTACGGGAGGCAGCAG-3’)和806R(5’-GGACTACHVGGGTWTCTAAT-3’)为引物扩增16S V3~V4区[13],采用MiSeq平台测序。
1.4 数据分析
测序数据经检测合格进行生物信息学分析。对有效数据在97%水平上进行操作分类单元(operational taxonomic units,OTU)聚类,并利用Greengenes数据库对OTU进行分类信息分析,分析样品物种组成[14-15]。利用Mothur软件进行Alpha多样性和Beta多样性分析。Beta多样性分析基于加权Unifrac算法进行主成分分析(principal component analysis,PCA)[16]。
2.1 3 种食用菌货架期内外源细菌数量变化
图1 3 种食用菌货架期外源细菌数量变化
Fig. 1 Changes in total number of exogenous bacteria in three kinds of mushrooms during storage
结果表明,货架期1~5 d内,3 种食用菌的外源细菌持续增殖,外源细菌数明显增加(图1)。金针菇外源细菌数量均明显高于杏鲍菇和蟹味菇,第5天可达到1.8×108CFU/g。在杏鲍菇货架期1 d时,外源细菌最少,货架期1~3 d,外源细菌增殖最快。蟹味菇在货架期外源细菌增殖相对比较缓慢。存在差异的主要原因可能是两方面:一是存在的外源细菌种类不一样;二是不同种类食用菌的质地和营养不同,为外源微生物生长繁殖创造的营养基础不同。
2.2 3 种食用菌货架期内外源细菌种类及变化
2.2.1 16S rDNA高通量测序细菌物种组成分析
对金针菇、杏鲍菇、蟹味菇1~5 d货架期外源细菌16S rDNA高通量测序,结果显示鉴定到31 个门,79 个纲,132 个目,212 个科,343 个属。
在不同分类级别,对外源细菌群落结构进行分析。门水平所有样品中含量最高是变形菌门(Proteobacteria)(84.7%),其次是厚壁菌门(Firmicutes)(9.4%)和拟杆菌门(Bacteroidetes)(4.4%)。纲水平所有样品含量最高的是γ-变形菌纲(Gammaproteobacteria)(78.4%),其次是杆菌纲(Bacilli)(8.8%)、α-变形菌纲(Alphaproteobacteria)(4.4%)、鞘脂杆菌纲(Sphingobacteria)(2.9%)。进一步在属水平进行分析,金针菇外源细菌中比例前10的属为乳球菌属(Lactococcus)、假单胞菌属(Pseudomonas)、沙雷氏菌(Serratia)、不动杆菌属(Acinetobacter)、明串珠菌属(Leuconostoc)、金黄杆菌属(Chryseobacterium)、肠杆菌属(Enterobacter)、爱文氏菌属(Ewingella)、欧文氏菌属(Erwinia)、柠檬酸杆菌属(Citrobacter)。杏鲍菇为假单胞菌属、乳球菌属、寡养单胞菌属(Stenotrophomonas)、芽孢杆菌属(Bacillus)、拟杆菌属(Bacteroides)、栖霉菌属(Mycetocola)、乳杆菌属(Lactobacillus)、鞘氨醇杆菌属(Sphingobacterium)、弓形杆菌属(Arcobacter)、苍白杆菌属(Ochrobactrum)。蟹味菇为假单胞菌属、寡养单胞菌属、鞘氨醇杆菌属、芽孢杆菌属、柠檬酸杆菌属、橄榄形菌属(Olivibacter)、欧文氏菌属、德沃斯氏菌(Devosia)、地杆菌属(Pedobacter)、弓形杆菌属。

图2 3 种食用菌货架期外源假单胞菌属及乳球菌属相对丰度变化
Fig. 2 Changes in relative abundance of exogenous Pseudomonas and Lactococcus in three kinds of mushrooms during storage
a.假单胞菌属相对丰度;b.乳球菌属相对丰度。
在属水平,假单胞菌属和乳球菌属丰度较高,在3 种食用菌表面均存在。蟹味菇外源细菌中假单胞菌属相对丰度远高于金针菇以及杏鲍菇;该属在杏鲍菇货架期延长过程中逐渐降低;在金针菇中相对丰度最低(图2a)。乳球菌属在不同货架期菌群结构中的变化与假单胞菌属有明显的区别。金针菇外源细菌中乳球菌属相对丰度最高,但随货架期延长丰度降低;杏鲍菇和金针菇呈现相同的变化趋势;蟹味菇在货架期乳球菌属丰度没有明显变化(图2b)。出现这样的现象可能是因为在货架期初期,富足的营养使菌体大量繁殖,伴随着新陈代谢产物的不断积累(如乳酸的积累),抑制细菌增殖,而部分能够适应环境改变的种类仍能大量存在(如假单胞菌)。
2.2.2 平板分离法物种鉴定
为进一步在种的分类水平进行分析,对金针菇、杏鲍菇、蟹味菇货架期1~5 d外源细菌进行平板分离、纯化和鉴定,共得到24 个属,38 个种(表1)。
表1 平板培养3 种食用菌分离外源细菌的种类
Table 1 Exogenous bacterial species in three kinds of mushrooms evaluated by plate culture method

注:+.有检出。
2.2.3 16S rDNA高通量测序和平板分离法物种鉴定结果对比
16S rDNA高通量测序发现金针菇、杏鲍菇、蟹味菇货架期外源细菌群落结构中相对丰度最高的属分别为乳球菌属、假单胞菌属、假单胞菌属;而平板培养法中,金针菇外源细菌中优势种为美洲爱文菌(爱文氏菌属)和变形肥杆菌(肥杆菌属),杏鲍菇为美洲爱文菌(爱文氏菌属)和北京泛菌(泛菌属),蟹味菇为解鸟氨酸拉乌尔菌(拉乌尔菌属)和深红沙雷氏菌(沙雷氏菌属)。
2 个方法存在一定的差异,但在平板培养法中检测到的优势种,均可在高通量测序方法中检测到。高通量测序法能更准确判读外源细菌群落种类相对丰度,且能够检测到外源细菌群落结构中更多的细菌种类,包括大量的不可培养细菌,但在种的分类水平鉴定方面并不准确,而平板培养法能准确进行种水平的分析。因此,将2 种方法相结合,能够更加科学地进行食用菌货架期外源细菌种类的分析。
2.3 3 种食用菌货架期外源细菌群落多样性分析
2.3.1 Alpha多样性分析
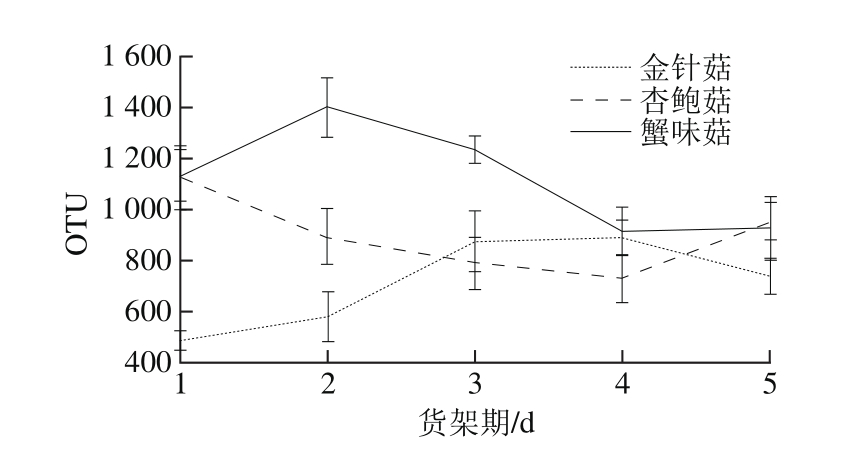
图3 3 种食用菌货架期外源细菌群落OTU数量变化
Fig. 3 Variations in OTU numbers of exogenous bacterial community in three kinds of mushrooms during storage
高通量测序结果显示,在0.97的相似度下,共获得8 480 个OTU。蟹味菇外源细菌OTU数量明显高于杏鲍菇和金针菇(图3)。金针菇外源细菌OTU数量在货架期1~4 d呈增长趋势,第5天部分下降;杏鲍菇外源细菌货架期OTU数量变化趋势与金针菇相反;蟹味菇外源细菌OTU数量在货架期1~2 d上升而后下降。
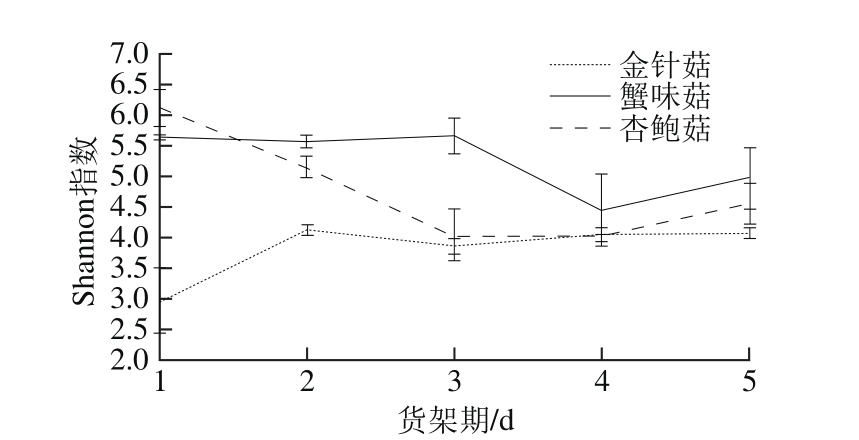
图4 3 种食用菌货架期外源细菌群落Shannon指数变化
Fig. 4 Variations in Shannon index of exogenous bacterial community in three kinds of mushrooms during storage
基于OTU的结果计算Shannon指数,结果表明在整个货架期内金针菇外源细菌群落多样性明显低于杏鲍菇和蟹味菇(图4)。杏鲍菇货架期内外源细菌Shannon指数呈现先下降后上升的变化,货架期1 d Shannon指数值最高;蟹味菇1~3 d货架期内外源细菌Shannon指数维持稳定,3~4 d转而降低,4~5 d有所回升。
2.3.2 Beta多样性分析
基于OTU的结果进行PCA分析,以观察不同食用菌不同货架期外源细菌群落间的差异(图5)。结果表明货架期1~5 d,金针菇、蟹味菇、杏鲍菇3 组样品内呈现一定的交叠现象,说明组内样品在货架期群落结构具有一定的相似性。PC1或PC2水平,金针菇细菌群落均能与杏鲍菇和蟹味菇区分开,说明该组样品相对于另外两组样品具有独特的群落结构。而杏鲍菇与蟹味菇这两组样品间菌群结构具有相似性。
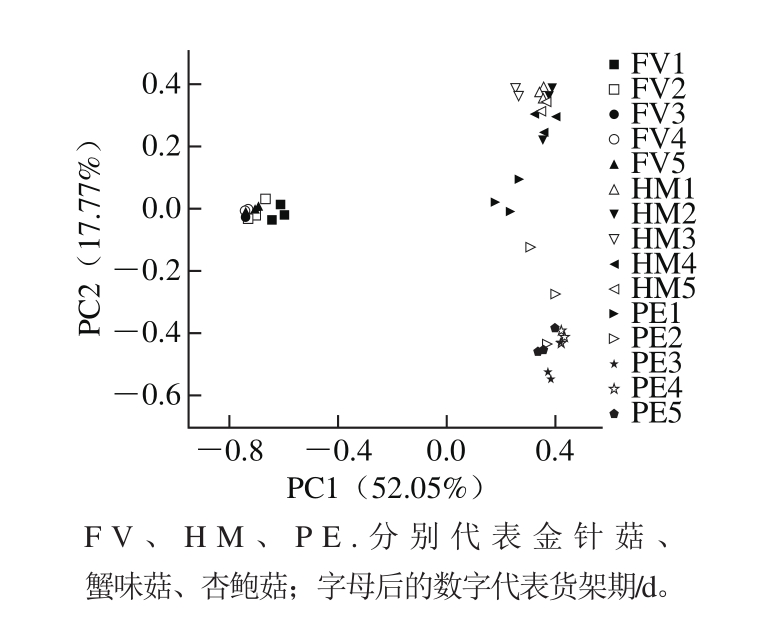
图5 3 种食用菌货架期外源细菌群落结构PCA图
Fig. 5 PCA plot for exogenous bacterial communities in three kinds of mushrooms stored for different time periods
在食品腐败过程中,微生物的作用是主要因素之一。以乳制品、肉制品、水产品和果蔬4 类食品为例,细菌的群体作用在这4 类食品腐败过程中起到关键作用[17]。同时有文献报道,细菌是污染鲜切蔬菜的主要微生物[18],如胡萝卜、紫甘蓝、西兰花的主要腐败细菌数量分别占总活菌数量的69%~75%、66%~70%、72%~77%。因此本研究中只探究了货架期外源细菌数量、种类、多样性等特征,而未对真菌群落进行研究。另外,鉴于低温贮藏可以减弱微生物繁殖,使肠杆菌、乳酸菌、假单胞菌等细菌的数量大幅降低,进而延长货架期,因此本研究只对货架期内(5 d)常温条件下贮藏的食用菌鲜品进行了研究。
本研究表明,不同种类食用菌外源细菌OTU组成存在较大差异,出现这样的情况可能是因为:1)不同食用菌种类质地与营养基础不同,因此为外源细菌提供的生长繁殖基础不同;2)不同食用菌种类子实体结构不同,与外源细菌接触机会不同;3)在3 种食用菌表面生存的细菌种类不同,有些种类生长繁殖能力强,在货架期能够一直繁殖,不会因为其他细菌的竞争关系而导致其数量的减少[19]。
鉴于食品安全考虑,外源细菌的致病性和致腐性值得关注,研究发现3 种食用菌外源细菌中存在部分条件致病菌及腐败菌,条件致病菌有腐生葡萄球菌[20]、木糖氧化无色杆菌[21]、不动杆菌属细菌[22]、金黄杆菌属细菌[23]、以及苍白杆菌属细菌[24]。同时在杏鲍菇、蟹味菇外源细菌中存在Arcobacter,一种人畜共患的病原菌,与人类胃肠炎和菌血症有关[25]。腐败细菌主要有美洲爱文菌[26]、拉恩氏菌属细菌[27]、假单胞菌属细菌[28-29]、泛菌属细菌[30]、乳球菌属细菌[31]、肠杆菌属细菌[31]以及欧文氏菌属细菌[31]。有报道在杏鲍菇子实体腐败组织中分离到泛菌属细菌[30],本实验同样在杏鲍菇中分离到;此外,有报道称美洲爱文菌是双孢蘑菇柄坏死的主要病原菌。从食品安全角度讲,应当对食用菌鲜品经过高温处理,减少外源细菌数量,避免有害细菌对人体的伤害。
参考文献:
[1] 郭乾元. 湖北省中小城市居民食用菌消费心理与消费行为研究[D].武汉: 华中农业大学, 2013: 17.
[2] 王琼英, 高巍, 陈强, 等. 与食用菌相关的食源性致病细菌简述[J]. 中国食用菌, 2016, 35(1): 59-61. DOI:10.13629/j.cnki.53-1054.2016.01.016.
[3] AGUIRRE L, FRIAS J M, BARRY-RYAN C, et al. Modelling browning and brown spotting of mushrooms (Agaricus bisporus)stored incontrolled environmental conditions using image analysis[J].Journal of Food Engineering, 2009, 91(2): 280-286. DOI:10.1016/j.jfoodeng.2008.09.004.
[4] 陈长宏, 张科, 陈环. 食品的细菌污染及预防[J]. 现代农业科技,2010(20): 348-350.
[5] 易琳琳, 应铁进. 食用菌采后品质劣变相关的生理生化变化[J].食品工业科技, 2012, 33(24): 434-436; 441. DOI:10.13386/j.issn1002-0306.2012.24.066.
[6] REINARTZ M, ALTER T, HILDEBRANDT G, et al. Impact of the deep freezing process and frozen storage on the microflora of two frozen convenience food products[J]. Fleischwirtschaft, 2011, 91(8):111-114.
[7] LEFF J W, NOAH F. Bacterial communities associated with the surfaces of fresh fruits and vegetables[J]. PLoS ONE, 2013, 8(3): 1-9.DOI:10.1371/journal.pone.0059310.
[8] JAWORSKA G, POGOŃ K, BERNAŚ E, et al. Vitamins, phenolics and antioxidant activity of culinary prepared Suillus luteus (L.) Roussel mushroom[J]. LWT-Food Science and Technology, 2014, 59(2):701-706. DOI:10.1016/j.lwt.2014.07.040.
[9] 卫生部. 食品微生物学检验 总则: GB 4789.1—2016[S]. 北京: 中国标准出版社, 2016: 1-5.
[10] 卫生部. 食品微生物学检验 菌落总数测定: GB 4789.2—2016[S].北京: 中国标准出版社, 2016: 1-5.
[11] FRANK J A, REICH C I, SHARMA S, et al. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes[J]. Applied Environmental Microbiology, 2008, 74(8):2461-2470. DOI:10.1128/AEM.02272-07.
[12] RUAN Z Y, ZHOU S, JIANG S H, et al. Isolation and characterization of a novel cinosulfuron degrading Kurthia sp. from a methanogenic microbial consortium[J]. Bioresource Technology, 2013, 147(9):477-483. DOI:10.1016/j.biortech.2013.08.017.
[13] DENNIS K L, WANG Y, BLATNER N R, et al. Adenomatous polyps are driven by microbe-instigated focal inflammation and are controlled by IL-10 producing T-cells[J]. Cancer Research, 2013, 73(19):5905-5913. DOI:10.1158/0008-5472.CAN-13-1511.
[14] DESANTIS T Z, HUGENHOLTZ P, LARSEN N, et al. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB[J]. Applied Environmental Microbiology, 2006, 72(7):5069-5072. DOI:10.1128/AEM.03006-05.
[15] WANG Q, GARRITY G M, TIEDJE J M, et al. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy[J]. Applied Environmental Microbiology, 2007,73(16): 5261-5267. DOI:10.1128/AEM.00062-07.
[16] FU Y J, LI X, ZHENG S H, et al. Classification and identification of bacteria in the soil treated by AcMNPV using high-throughput sequencing technique[J]. Biotechnology and Bioprocess Engineering,2015, 20(5): 931-936. DOI:10.1128/AEM.00062-07.
[17] 李学鹏, 陈桂芳, 仪淑敏, 等. 食品腐败中细菌群体感应现象的研究进展[J]. 食品与发酵工业, 2015, 41(8): 244-250. DOI:10.13995/j.cnki.11-1802/ts.201508046.
[18] 陈湘宁, 钟思琼, 金文斌, 等. 膜包装鲜切蔬菜中主要腐败菌的分离与鉴定[J]. 中国食品学报, 2012, 12(5): 154-160. DOI:10.16429/j.1009-7848.2012.05.028.
[19] 姜天甲. 主要食用菌采后品质劣变机理及调控技术研究[D]. 杭州:浙江大学, 2010: 53.
[20] 幸雄飞. 泌尿生殖道腐生葡萄球菌的感染及临床意义[J]. 现代医院,2006, 6(8): 68-69.
[21] 李金钟. 木糖氧化无色杆菌的研究进展[J]. 临床检验杂志, 2009,27(1): 72-73. DOI:10.13602/j.cnki.jcls.2009.01.021.
[22] CARVALHEIRA A, FERREIRA V, SILVA J, et al. Enrichment of Acinetobacter spp. from food samples[J]. Food Microbiology, 2016,55: 123-127. DOI:10.1016/j.fm.2015.11.002.
[23] 陈吉泉, 颜泽敏. 金黄杆菌致下呼吸道感染的病原学和临床研究[J].中华医院感染学杂志, 2003, 13(10): 975-977. DOI:10.3321/j.issn:1005-4529.2003.10.033.
[24] 刘志国, 崔步云, 夏咸柱. 人苍白杆菌研究进展[J]. 微生物学报,2015, 55(8): 977-982. DOI:10.13343/j.cnki.wsxb.20140534.
[25] 毕水莲, 孟赫诚. 致病性弓形杆菌属生物学特性及诊断研究进展[J]. 现代食品科技, 2013, 29(1): 211-214. DOI:10.13982/j.mfst.1673-9078.2013.01.030.
[26] MADBOULY A K, EL-SHATOURY E H, ABOUZEID M A, et al.Etiology of stipe necrosis of cultivated mushrooms (Agaricus bisporus)in Egypt[J]. Phytopathologia Mediterranea, 2014, 53(1): 124-129.DOI:10.14601/Phytopathol_Mediterr-13224.
[27] ONGENG D, DEVLIEGHERE F, DEBEVERE J, et al. The efficacy of electrolysed oxidising water for inactivating spoilage microorganisms in process water and on minimally processed vegetables[J].International Journal of Food Microbiology, 2006, 109(3): 187-197.DOI:10.1016/k.ijfoodmicro.2005.12.013.
[28] PINTO L, IPPOLITO A, BARUZZI F. Control of spoiler Pseudomonas spp. on fresh cut vegetables by neutral electrolyzed water[J]. Food Microbiology, 2015, 50: 102-108. DOI:10.1016/j.fm.2015.04.003.
[29] FRANZETTI L, SCARPELLINI M. Characterisation of Pseudomonas spp. isolated from foods[J]. Annals of Microbiology, 2007, 57(1): 39-47. DOI:10.1016/j.fm.2015.04.003.
[30] LIU Y, WANG S X, ZHANG D P, et al. Pantoea beijingensis sp. nov.isolated from the fruiting body of Pleurotus eryngii[J]. Antonie van Leeuwenhoek, 2013, 104(6): 1039-1047. DOI:10.1007/s10482-013-0024-0.
[31] NGUYEN-THE C, CARLIN F. The microbiology of minimally processed fresh fruits and vegetables[J]. Critical Reviews in Food Science and Nutrition, 1994, 34(4): 371-401.DOI:10.1080/10408399409527668.
Dynamic Changes of Exogenous Bacteria in Marketed Fresh Mushrooms
WANG Qiongying, GAO Wei, ZHAO Mengran, WU Xiangli, CHEN Qiang, QU Jibin, HUANG Chenyang*
(Key Laboratory of Microbial Resources Collection and Preservation, Ministry of Agriculture, Institute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences, Beijing 100081, China)
Abstract:Edible mushrooms are popular with consumers due to their rich nutrition, tasty and health benefits. Fresh edible mushrooms are usually placed at room temperature in the market. Fresh mushrooms are readily infected with exogenous microbes for their rich nutrition, tender surface structure and lack of a protective structure. In this paper, fresh mushrooms of Flammulina valutipes, Pleurotus eryngii, and Hypsizygus marmoreus were studied. The exogenous bacterial number, species and community diversity were analyzed as a function of storage time at ambient temperature. Results showed that the amount of exogenous bacteria in F. valutipes, P. eryngii and H. marmoreus increased significantly by 29, 417 and 4 116 times,respectively. Besides, the abundance of different exogenous bacteria also changed during the storage period of 5 days, with the highest increase of Pseudomonas spp. in H. marmoreus and decline of Lactococcus spp. in F. valutipe being observed.According to Shannon index, exogenous bacterial diversity of F. valutipe was significantly lower than that of P. eryngii, and H. marmoreus. Principal component analysis results showed that the microbial community structure was similar at different times of storage within each species, and H. marmoreus and P. eryngii showed some similarities, while differences between F. velutipes and the other two species were observed.
Key words:storage period; high throughput sequencing; diversity; pathogenic bacteria; spoilage bacteria
DOI:10.7506/spkx1002-6630-201721013
中图分类号:S646
文献标志码:A
文章编号:1002-6630(2017)21-0080-06
引文格式:王琼英, 高巍, 赵梦然, 等. 市售食用菌鲜品外源细菌的动态变化分析[J]. 食品科学, 2017, 38(21): 80-85.
DOI:10.7506/spkx1002-6630-201721013. http://www.spkx.net.cn
WANG Qiongying, GAO Wei, ZHAO Mengran, et al. Dynamic changes of exogenous bacteria in marketed fresh mushrooms[J]. Food Science, 2017, 38(21): 80-85. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201721013. http://www.spkx.net.cn
收稿日期:2017-03-09
基金项目:“十二五”国家科技支撑计划项目(2013BAD16B03)
作者简介:王琼英(1993—),女,硕士研究生,研究方向为食用菌遗传育种。E-mail:735665861@qq.com
*通信作者:黄晨阳(1977—),男,副研究员,博士,研究方向为食用菌遗传育种。E-mail:huangchenyang@caas.cn