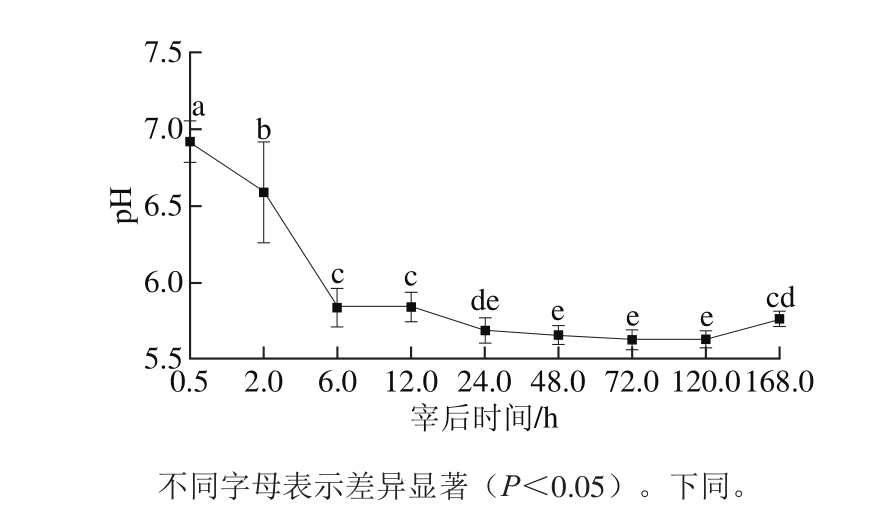
图1 宰后不同时间pH值的变化
Fig. 1 Variation in pH during 168.0 h post-mortem
李桂霞1,2,李 欣2,李 铮2,王 颖1,2,朱 杰1,*,张德权2,*
(1.西北农林科技大学理学院,生物物理研究所生物力学与工程研究室,陕西 杨凌 712100;2.中国农业科学院农产品加工研究所,农业部农产品加工重点实验室,北京 100193)
摘 要:动物宰后从肌肉到肉品经过僵直、成熟等一系列复杂的生理生化反应。僵直及成熟过程的研究可为肉品质改善及肉制品加工提供理论依据。选取杂交公羊(小尾寒羊×北京本地羊)双侧背最长肌在4 ℃成熟0.5、2.0、6.0、12.0、24.0、48.0、72.0、120.0、168.0 h,测定不同时间的pH值、剪切力、肌节长度、ATP含量、肌原纤维小片化指数(myofibril fragmentation index,MFI)、钙蛋白酶活力。结果表明:宰后成熟过程中,pH值先下降后逐渐趋于稳定,从第24小时后变化不显著(P>0.05);剪切力先上升后下降,在第24小时达到最大值;肌节长度在宰后先缩短后逐渐变长,且在第48小时缩至最短;ATP含量先上升后下降,48 h后趋于稳定;μ-钙蛋白酶80 kD大亚基在宰后24 h基本降解完全,其降解的78 kD大亚基在第48小时降解完全;肌间线蛋白和肌钙蛋白T作为μ-钙蛋白酶的降解底物,在成熟过程中发生降解,第168小时几乎观察不到完整的蛋白条带;随着宰后时间的延长,羊背最长肌从第2小时开始僵直,到第24小时程度达到最大,从第48小时开始解僵,解僵后肉的嫩度逐渐改善。
关键词:羊背最长肌;僵直;嫩度;成熟
僵直是宰后肌肉必经阶段,是体内能量消耗的体现。解僵过程对肉嫩度的改善有很大影响,所以对僵直及成熟进程的研究可为肉品质改善及肉制品加工提供理论依据。僵直进程受许多因素影响,主要包括年龄、性别、胶原蛋白含量、糖酵解速率、肌肉收缩和肌纤维蛋白降解等。因此,宰后肌肉所发生的各种反应与活体肌肉完全处于不同状态、进行着不同性质的反应,研究这些特性对于了解宰后不同僵直阶段以及成熟过程中肉的性质变化,改善肉的品质以及指导肉制品加工有着重要意义。
畜禽屠宰后由于血液循环和呼吸终止,而细胞仍然进行着代谢活动,细胞内很快变成无氧状态,肌肉开始僵直。僵直是肌肉转变为肉品过程中发生的最显著变化[1]。僵直过程主要分为3 个阶段,即僵直迟滞期、僵直急速形成期和僵直后期[2-3]。畜禽宰后早期发生僵直,肉的硬度逐渐增加、嫩度变差,在僵直结束时嫩度最小,而在随后的解僵成熟过程中肉的嫩度逐渐得到改善。大量研究表明,在宰后成熟过程中,Ca2+的释放、肌动球蛋白解离、肌原纤维蛋白降解以及蛋白酶水解活性对肉的嫩度起到积极的作用[2-6]。张丽等[7]在研究牦牛肉品质时,提出延长成熟期来提高肉的嫩度。Wu Gaojie等[8]发现肌原纤维蛋白的快速降解是肉嫩度改善的关键因素。贾小翠等[9]研究发现,禁食组以僵直后的肉加工的鸡肉肠质构特性劣于以僵直和僵直前的肉加工的鸡肉肠,僵直前制成的鸡肉肠表现出较高的持水力和较好的弹性、黏聚性和胶黏性,硬度和咀嚼性稍高,但总体上未禁食组僵直前制成的肉糜制品加工特性较好,表明不同僵直阶段肉的加工特性存在差异。因此,了解肉品在宰后不同僵直阶段和成熟过程的特性对于肉品质改善和肉制品加工有指导意义。
本研究采用杂交公羊(小尾寒羊×北京本地羊)的背最长肌为原料,分析了宰后不同时间pH值、ATP含量、肌节长度、剪切力、肌原纤维小片化指数(myofibril fragmentation index,MFI)、钙蛋白酶活性以及肌间线蛋白和肌钙蛋白T的降解程度,明确了宰后羊肉在不同时间的僵直程度以及成熟过程,可为预测和控制宰后肌肉嫩化的速度和程度提供理论依据。
1.1 材料与试剂
选用12 月龄、胴体质量25 kg左右的6 只杂交公羊(小尾寒羊×北京本地羊),未去势,清真屠宰放血开始计时,宰后0.5 h后迅速取出两侧的背最长肌,装入标记好的自封袋,置于冰盒中并在2 h内运回实验室,将样品取出用保鲜膜包装于4 ℃保藏。
蛋白酶抑制剂 瑞士Roche公司;蛋白浓度测定试剂盒 美国Pierce公司;肌间线蛋白单克隆抗体(D1033)、二抗(辣根过氧化酶标记的羊抗鼠免疫球蛋白G)、三羟甲基氨基甲烷(tris base,Tris)、十二烷基硫酸钠(sodium dodecyl sulfate,SDS)、丙烯酰胺、甲叉双丙烯酰胺、过硫酸铵、四甲基乙二胺(tetramethylethylenediamine,TEMED) 美国Sigma公司;ATP含量测试盒 南京建成生物工程研究所;肌钙蛋白T单克隆抗体(ab130003) 英国Abcam公司;乙醇、乙酸等(均为分析纯) 北京化学试剂公司。
1.2 仪器与设备
205便携式pH计 德国德图公司;TA-XT2i质构仪英国Stable Micro Systems公司;Ultra Turrax Disperser S25分散器 德国IKA公司;Neofuge高效冷冻离心机上海力申科学仪器有限公司;UV-1800紫外-可见分光光度计 岛津仪器(苏州)有限公司;ML204/02电子天平 梅特勒-托利多(上海)有限公司;Chameleon V多功能酶标仪 芬兰Hidex公司;电泳设备(Mini-PROTEAN Tetra System)、全自动半干转印仪(Trans-Blot Turbo System) 美国Bio-Rad公司;Typhoon Trio多功能激光成像系统 美国GE公司;HH-4数显恒温水浴锅 江苏省金坛市荣华仪器制造有限公司;MJ-Ⅱ霉菌培养箱 上海一恒科技有限公司。
1.3 方法
1.3.1 样品处理
羊背最长肌于宰后0.5、2.0、6.0、12.0、24.0、48.0、72.0、120.0、168.0 h测定pH值;于2.0、12.0、24.0、48.0、72.0、120.0 h测定剪切力;并在各时间点取2 mm×2 mm×5 mm的细条置于戊二醛中,一部分用于测定肌节长度,另一部分液氮速冻,于-80 ℃贮藏备用以测定ATP含量、MFI、μ-钙蛋白酶活力和肌原纤维蛋白的变化。
1.3.2 pH值测定
采用便携式pH计插入肌肉2 cm深处,避开脂肪和筋膜。连续测定3 次,结果取平均值。
1.3.3 ATP含量测定
称取1 g样品,加入9 mL的沸水并匀浆1 min,之后在沸水中煮10 min,3 000×g离心10 min,可得到质量分数为10%的组织匀浆液,再用双蒸水稀释20 倍,获得质量分数为0.5%的组织匀浆液,从其中取30 μL,采用ATP含量测试盒测定羊肉宰后0.5、2.0、6.0、12.0、24.0、48.0、72.0、120.0、168.0 h ATP含量的变化,使用二喹啉甲酸(bicinchoninic acid,BCA)试剂盒测定蛋白质量浓度。以每克蛋白含有ATP的物质的量作为ATP含量单位。
1.3.4 剪切力测定
用质构仪测定剪切力。取体积约3 cm×6 cm×6 cm肉块除去表面的脂肪和筋膜,装入蒸煮袋中,将温湿度记录仪的探头沿肌纤维方向插入肉的中心,置于水浴锅在80 ℃温度条件下加热至中心温度为70 ℃后立即取出,擦去表面渗出的水分,待温度降到室温,沿肌纤维方向切成1.0 cm×1.0 cm×1.5 cm的肉块。
质构仪测定条件:探头型号为HDP/BSW探头,测前速率1.0 mm/s,测中速率1.0 mm/s,测后速率5.0 mm/s,时间间隔5 s。每个样品7~10 次重复,取平均值。
1.3.5 肌原纤维超微结构测定
参考王欣[10]和宋洁[11]等的方法并作修改。宰后立即取羊背最长肌,在不同宰后时间沿肌纤维方向切成1 mm×1 mm×3 mm规格的肉条,取样时避免牵拉和钳夹,以免增大实验误差,放入体积分数为2.5%的戊二醛溶液中,在室温(25℃)条件下固定。室温条件下用0.1 mol/L的磷酸盐缓冲液(pH 7.3)冲洗,质量分数为1%的四氧化锇固定,放置2 h,磷酸盐缓冲液冲洗后,采用30%、50%、70%、85%、96%的乙醇溶液和无水乙醇梯度脱水以及无水丙酮置换3次。树脂包埋完放入烘箱进行聚合,采用超薄切片机切片,醋酸铀-柠檬酸铅双染色,透射电镜观察拍照。Image-Pro Plus 6.0软件分析透射电子显微镜图片,测量肌节长度。
1.3.6 MFI测定
参照Culler等[12]的方法并稍作调整。称取0.5 g肉样,切碎后加入5 mL预冷的MFI缓冲液(100 mmol/L KCl、20 mmol/L K2HPO4、1 mmol/L乙二胺四乙酸、1 mmol/L MgCl2),高速匀浆3 次,每次30 s,中间间隔1 min。于3 000×g、4 ℃条件下离心15 min,去除上清液,将沉淀用5 mL预冷的MFI缓冲液悬浮,在3 000×g、4℃条件下再次离心15 min,弃去上清液,用1.25 mL预冷的MFI缓冲液将沉淀充分悬浮。将悬浮液用20 目筛网过滤除去结缔组织,再加1.25 mL预冷的MFI缓冲液清洗离心管,使肌原纤维通过筛孔。过滤后的悬浮液用双缩脲法测定蛋白质量浓度,然后用MFI缓冲液调整悬浮液蛋白质量浓度为(0.50±0.05) mg/mL,在540 nm波长处测定吸光度,将所得结果乘200后即可得到MFI。
1.3.7 肌原纤维蛋白降解测定
肌钙蛋白T和肌间线蛋白的降解测定采用蛋白质免疫印迹技术。将1 g肌肉组织加入6 mL预冷的缓冲剂中(100 mmol/L Tris、10 mmol/L二硫苏糖醇、蛋白酶抑制剂(50 mL缓冲液中添加一片),pH 8.3,需现配现用),用Ultra Turrax T25匀浆机4档匀浆2 次,每次15 s,于-80 ℃温度条件下保存。于10 000×g,4 ℃离心35 min,所得沉淀即肌原纤维蛋白。沉淀溶解于5% SDS溶液(60 ℃),然后用匀浆机3档(约9 500 r/min)匀浆30 s,80 ℃加热20 min。
提取的肌原纤维蛋白样品调整至统一浓度后,按体积比为1∶1加入样品缓冲液(10% SDS、纯甘油、0.5 mol/L Tris-HCl(pH 6.8)、1 mol/L二硫苏糖醇、1 mol/L溴酚蓝),沸水浴中加热5 min,冷却后于12 000×g离心1 min,取上清液上样,上样量为5 μg。SDS-聚丙烯酰胺凝胶电泳中肌间线蛋白和肌钙蛋白T分别采用12%、15%的分离胶与4%的浓缩胶(质量比为37.5∶1.0的丙烯酰胺和甲叉双丙烯酰胺混合溶液)。电泳初始电压为70 V,待蛋白进入分离胶后调整电压为110 V,至蛋白到达离凝胶底部5 mm处停止电泳。肌钙蛋白T采用湿法将凝胶上的蛋白条带转移至聚偏二氟乙烯(polyvinylidene fluoride,PVDF)膜上,100 V恒压条件下,冰浴转膜60 min;肌间线蛋白采用半干法,在全自动半干转印仪中,于25 V、2.5 A条件下转膜3 min。转膜结束后,将PVDF膜在TBS缓冲液(0.01 mol/L Tris、0.15 mol/L NaCl,pH 7.5)中清洗3 次,每次1 min,在封闭液(含0.05%吐温20、3%牛血清白蛋白的TBS溶液)中室温封闭2 h。
一抗使用肌间线蛋白和肌钙蛋白T单克隆抗体,稀释1 000 倍,4 ℃过夜孵育;随后将膜用TBST1溶液(含0.1%吐温20的TBS溶液)漂洗3 次后与二抗(稀释2 500倍)室温孵育2 h;孵育结束后用TBST2溶液(0.05 mol/L Tris、0.15 mol/L NaCl、0.1%吐温20,pH 7.5)漂洗PVDF膜3 次,每次10 min,电化学发光(electro-chemi-luminescence,ECL)法显色曝光,并使用凝胶成像仪拍照。
1.3.8 钙蛋白酶活力的测定
采用活性电泳的方法进行测定,参考Kadee[13]和Melody[14]等的方法并适当调整。取0.5 g样品,加入3 倍体积的抽提液(100 mmol/L Tris、10 mmol/L 乙二胺四乙酸、0.05%巯基乙醇,pH 8.3),进行匀浆,然后于10 000×g 4 ℃离心35 min,上清液为所需蛋白样品,采用BCA法测定蛋白质量浓度,并调整至统一质量浓度。按体积比为3∶2加入样品缓冲液(150 mmol/L Tris-HCl(pH 6.8)、20%甘油、0.75%巯基乙醇、质量分数0.02%溴酚蓝),混匀后备用。采用12.5%的酪蛋白分离胶(质量比75∶1的丙烯酰胺与甲叉双丙烯酰胺混合溶液、375 mmol/L Tris-HCl(pH 8.8)、0.05 g/mL TEMED、0.05 g/mL过硫酸铵、2.1 mg/mL酪蛋白)和4%的浓缩胶(质量比为37.5∶1的丙烯酰胺和甲叉双丙烯酰胺混合溶液、125 mmol/L Tris-HCl(pH 6.8)、0.05 g/mL TEMED 、0.05 g/mL过硫酸铵,凝胶厚度为0.75 mm。上样前,在冰浴中,100 V恒压预电泳15 min,上样量为40 μg,在相同电泳缓冲液(25 mmol/L Tris-HCl、0.53 g/L巯基乙醇、192 mmol/L甘氨酸、1 mmol/L乙二胺四乙酸,pH 8.3)中冰浴,于100 V恒压条件下电泳7 h。电泳结束后更换3 次孵育缓冲液(50 mmol/L Tris、0.53 g/L巯基乙醇、4 mmol/L CaCl2,pH 7.5),每次室温孵育20 min,并在上述相同的缓冲液中室温孵育16 h。凝胶经考马斯亮蓝染色脱色后,使用凝胶成像系统照相。
1.4 统计分析
实验结果用SPSS 22.0和Excel 2010软件进行处理,同一处理组的不同时间点之间采用F检验进行差异显著性分析,采用Duncan多重比较法进行差异显著性分析,显著水平为P<0.05。
2.1 pH值
图1 宰后不同时间pH值的变化
Fig. 1 Variation in pH during 168.0 h post-mortem
pH值反映肌肉内乳酸和无机磷酸的沉积,间接反映糖酵解程度。由图1可知,随着宰后时间的延长,宰后24.0 h内pH值逐渐下降且差异显著(P<0.05),24.0 h后变化趋于稳定,说明糖酵解反应几乎终止,而在第168小时,pH值升高,可能是由于微生物繁殖或者蛋白质降解造成的。吴菊清等[15]采用了相同的方法研究牛肉和猪肉pH值变化,其变化趋势与本研究结果相似。
2.2 ATP含量
ATP是肌肉运动的直接能量载体,肌肉收缩过程中ATP的状态影响肌动蛋白和肌球蛋白的状态。肌球蛋白和肌动蛋白结合形成肌动球蛋白,结合力取决于与核苷酸的结合状态,未结合核苷酸的肌球蛋白与肌动蛋白的亲和力最高。随着宰后ATP的消耗,肌动蛋白和肌球蛋白之间形成强结合横桥,即僵直状态。
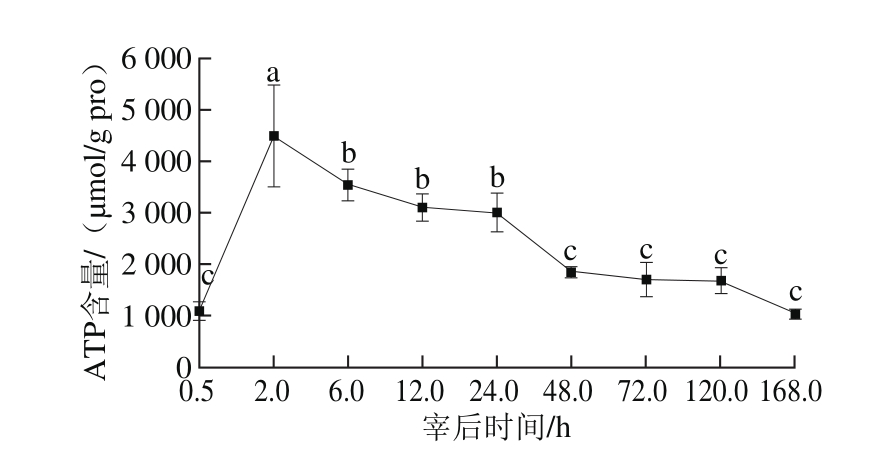
图2 宰后不同时间ATP含量的变化
Fig. 2 Variation in ATP content during 168.0 h post-mortem
由图2可知,ATP含量先增大后逐渐下降,可能是宰后胴体供氧停止,启动磷酸肌酸供能途径,使得ATP含量逐渐上升。肌肉细胞的磷酸肌酸含量是ATP的3~4 倍,可贮存供短期活动用的、足够的磷酸基团,在肌酸激酶的作用下很快供给ADP磷酸基,使之再合成ATP。磷酸肌酸耗尽时启动糖酵解途径,其产生速率迅速下降,而肌肉收缩需要不断的能量供给,ATP含量逐渐下降。Frylinck[16]和徐昶[17]等研究发现牛背最长肌ATP含量随宰后时间的延长而逐渐下降。到第48小时后ATP含量趋于稳定,可能是糖酵解反应终止,肌动球蛋白之间不再形成横桥,僵直达到最大。综上所述,ATP含量在宰后48.0h内显著下降(P<0.05),从第48小时开始ATP含量趋于稳定,说明僵直完成,推测解僵在宰后第2天发生。
2.3 肌原纤维的超微结构及肌节长度
随着宰后成熟时间的延长,肌原纤维的结构被破坏,Z线结构由原来的完整形态变得模糊,表明与Z线结构相关的蛋白发生降解;M线的结构相对稳定,从宰后0.5~168.0 h均可观察到(图3)。
如图4所示,宰后随着成熟时间的延长,肌节长度第48小时最短,从第48小时开始肌节逐渐变长[18],说明肌肉从第48小时开始解僵。宰后成熟过程中,肌肉经过尸僵、解僵的过程,在这个过程中,肌节长度先变短后变长,与Geesink等[19]的研究结果相似。在动物宰后初期,磷酸肌酸提供高能磷酸键形成ATP,耗尽后由糖酵解提供ATP,其含量迅速下降,肌动蛋白和肌球蛋白之间形成不可逆横桥,肌肉收缩至伸缩性几乎完全消失,肌节长度最短。之后肌节逐渐变长,可能是肌原纤维蛋白降解和肌动球蛋白解离的共同作用造成的[20]。
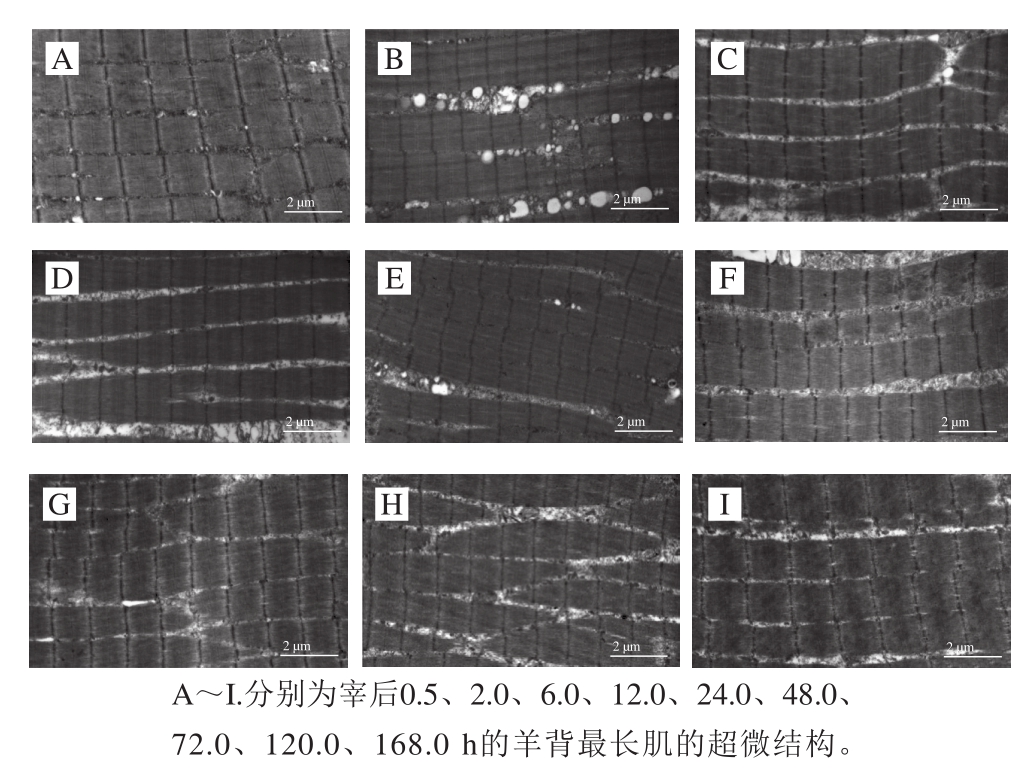
图3 宰后肌原纤维超微结构变化
Fig. 3 Variationin in the ultrastructure of myofibril during 168.0 h post-mortem
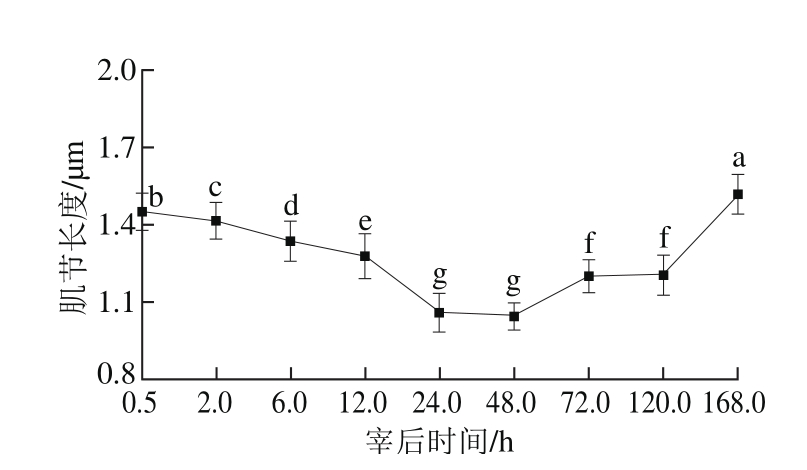
图4 宰后不同时间肌节长度的变化
Fig. 4 Variation in sarcomere length during 168.0 h post-mortem
2.4 剪切力
剪切力是衡量肌肉嫩度的一个重要指标,剪切力越大说明嫩度越差。宰后羊肉经历僵直、解僵和成熟的过程,剪切力在僵直开始后不断升高。随着宰后时间的延长,僵直解除,剪切力值开始下降,嫩度上升,肌肉成熟[21]。
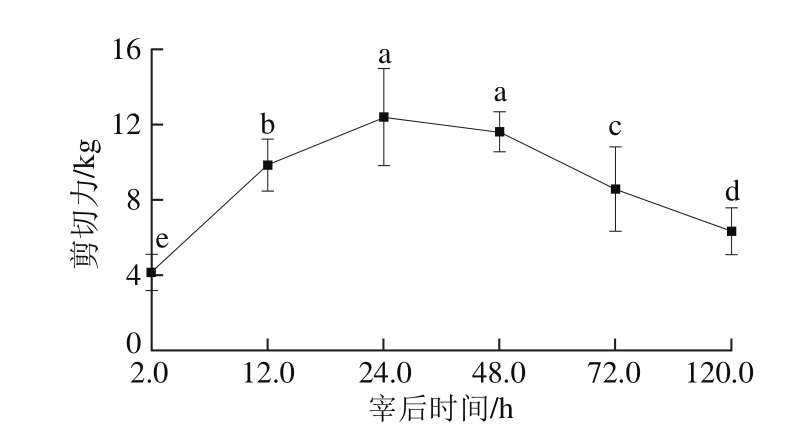
图5 宰后不同时间剪切力的变化
Fig. 5 Variation in shear force during 120.0 h post-mortem
如图5所示,随着宰后时间的延长,剪切力显著上升且在第24小时达到最大值,但是与宰后第48小时相比差异不显著(P>0.05),而肌节长度在宰后第24小时和第48小时较短,与剪切力结果相反,肌节长度越短,剪切力越大。表明僵直主要发生在宰后24.0 h内,经过一段僵直期在第48小时后开始解僵,剪切力逐渐减小,差异显著(P<0.05)。刘佳东[22]和Hopkins[23]等在研究牦牛肉排酸过程中剪切力的变化,其变化趋势和本研究是相似的,但是在第4天达到最大值,说明不同物种的僵直时间不同。
2.5 肌原纤维小片化指数
MFI是反映肌纤维内部肌原纤维及骨架蛋白完整程度的指标,μ-钙蛋白酶是导致MFI升高最主要的内源酶。MFI值越大,表明肌原纤维内部结构的完整性受到破坏的程度越大,嫩度越好。
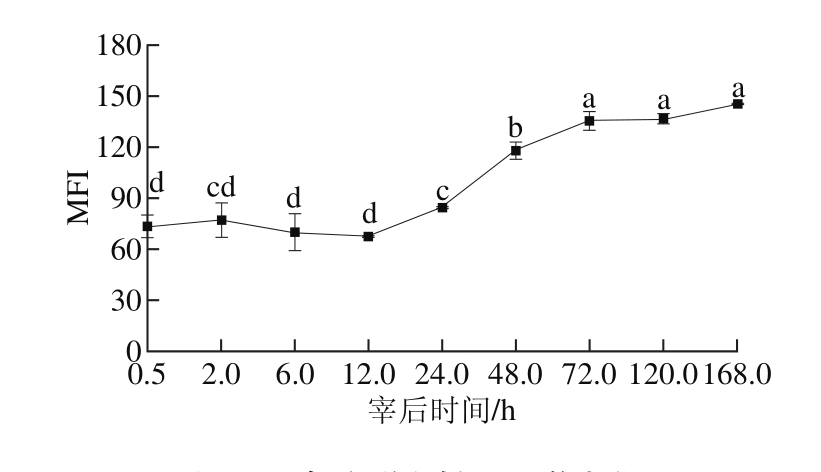
图6 宰后不同时间MFI的变化
Fig. 6 Variation in MFI during 168.0 h post-mortem
如图6所示,MFI在宰后12.0 h内变化差异不显著(P>0.05),随着宰后时间的延长,MFI值逐渐增大且在12.0~72.0 h显著上升(P<0.05),表明这期间肌原纤维骨架受到破坏,成熟过程可能主要完成于这段时间,第72小时以后MFI变化不显著(P>0.05),表明小片化程度趋于稳定。师希雄等[24]在研究牦牛肉宰后成熟过程中MFI的变化表明,整体变化趋势呈S形,与本实验的结果相似,李培迪[25]和李诚[26]等研究在冰温和冷藏贮存下羊背最长肌MFI的变化,也验证了同样的现象。
2.6 钙蛋白酶活力
钙蛋白酶分为μ-钙蛋白酶和m-钙蛋白酶两类,自溶后才表现活性,其活性下降是其发挥蛋白水解作用的标志,其中μ-钙蛋白酶是一种钙激活中性半胱氨酸内肽酶,主要分布在肌原纤维的Z盘附近[27-28]。体内μ-钙蛋白酶和m-钙蛋白酶都能水解肌原纤维蛋白,但在宰后成熟的过程中m-钙蛋白酶几乎没有水解活性,而μ-钙蛋白酶在自溶中发挥活性后失活。

图7 宰后不同时间钙蛋白酶活力电泳分析
Fig. 7 Casein zymography of calpain during 168.0 h post-mortem
由图7所示,随着宰后成熟时间的延长,m-钙蛋白酶的活力从0.5~168.0 h变化不大,表明在宰后成熟过程中没有发挥水解蛋白的作用;而μ-钙蛋白酶的大亚基在自溶过程中发挥活性,由80 kD降解为78 kD,最终降解为76 kD。本实验从宰后0.5 h开始,80 kD的大亚基活力逐渐下降,78 kD的大亚基活力先上升后下降,到第48小时活力完全消失。综上所述,μ-钙蛋白酶80 kD大亚基的初始含量高,但降解速度快,在宰后24.0 h失活,其降解的78 kD大亚基在48.0 h失活。对照上述结果,μ-钙蛋白酶失活的时间点是解僵开始的时间点,μ-钙蛋白酶发挥活力期间剪切力是上升的,表明其活力的发挥在宰后48.0 h内对僵直进程的影响作用不明显。
2.7 肌原纤维蛋白的降解分析
肌间线蛋白和肌钙蛋白T作为μ-钙蛋白酶的主要底物,其降解程度可反映肌原纤维蛋白整体降解程度。肌间线蛋白围绕Z盘分布并延伸到Z盘内部,是位于Z线以及Z线和肌细胞膜之间的骨架蛋白,其亚基的分子质量约53 kD,在成熟的肌细胞中则起着连接每条肌原纤维并将其与细胞膜连结,起着固定肌原纤维的作用,进而维持整个骨骼肌细胞的有序性和完整性等。肌钙蛋白T为肌细丝蛋白,主要结合原肌球蛋白,作为肉嫩度改善的一个标志,其降解破坏了肌细丝的完整性,改变了肌动蛋白和肌球蛋白之间的相互作用,导致了肌原纤维的片段化。
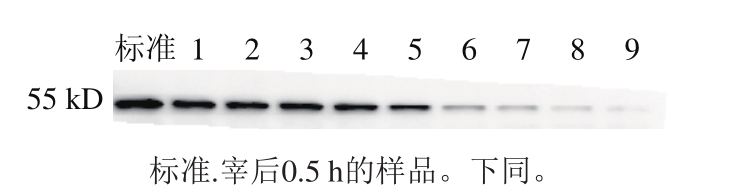
图8 宰后不同时间肌间线蛋白降解的免疫印迹图
Fig. 8 Representative Western-blot pattern of desmin degradation during 168.0 h post-mortem
由图8可知,随着宰后时间的延长,肌间线蛋白的相对含量逐渐减小,特别是从宰后第48小时开始相对含量降低,到第168小时几乎降解完全,表明Z线的结构受到破坏。Lomiwes等[29]也发现了同样的变化趋势。
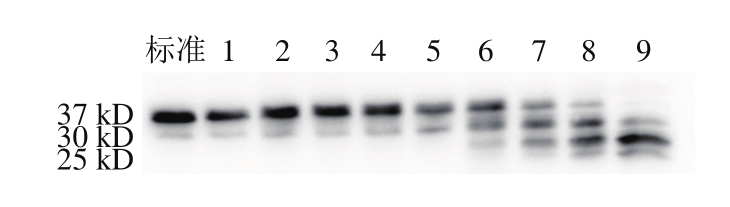
图9 宰后不同时间肌钙蛋白T的免疫印迹图
Fig. 9 Representative Western-blot profile of troponin T degradation during 168.0 h post-mortem
由图9可知,肌钙蛋白T在宰后随着时间的延长发生降解,由完整的37 kD降解为30 kD的蛋白,特别是从宰后24.0 h后,降解条带的相对含量逐渐增大,完整条带的相对含量逐渐减小至消失。Marino等[30]在研究不同品种牛背最长肌也得出了相似的结果。Kim等[31]研究报道冷冻/解冻后肌间线蛋白和肌钙蛋白T随着成熟时间的延长,降解程度逐渐增大。由μ-钙蛋白酶活力结果表明其蛋白水解活力主要在宰后48.0 h内发挥作用,而肌间线蛋白和肌钙蛋白T在第72小时后仍在降解,可能是肌原纤维蛋白的降解滞后于μ-钙蛋白酶发挥活性的时间或者其他蛋白酶(如组织蛋白酶)发挥水解蛋白活性[32]。肌原纤维蛋白的降解生成了小肽段,推测可能是碱性基团暴露增多,从而导致pH值上升。综上所述,宰后成熟过程中肌间线蛋白和肌钙蛋白T发生降解,从第48小时开始,蛋白的降解速率加快,说明解僵也从此时开始。
随着宰后时间的延长,羊背最长肌经历了从僵直到解僵的过程。pH值于宰后第24小时开始趋于稳定;在第72小时剪切力显著减小(P<0.05),肌节长度显著变长(P<0.05),所以宰后第72小时是羊背最长肌肉品质转好的关键转折。μ-钙蛋白酶活力在宰后48.0 h消失,而肌间线蛋白和肌钙蛋白T从第72小时开始大量下降,说明酶活力发挥和蛋白降解不同步。总之,羊背最长肌从宰后2.0 h开始进入僵直到24.0 h达到最大,经过一段僵直后期,从第48小时开始解僵,嫩度逐渐改善。
参考文献:
[1] 陈立娟, 李欣, 杨扬, 等. 不同嫩度羊肉肌浆蛋白质磷酸化水平随宰后成熟时间变化的研究[J]. 现代食品科技, 2015, 31(4): 95-101.DOI:10.13982/j.mfst.1673-9078.2015.4.016.
[2] 李兰会, 孙丰梅, 黄娟, 等. 宰后肉品pH值与嫩度[J]. 肉类工业,2006(12): 28-30. DOI:10.3969/j.issn.1008-5467.2006.12.012.
[3] WANG D Y, DONG H, ZHANG M H, et al. Changes in actomyosin dissociation and endogenous enzyme activities during heating and their relationship with duck meat tenderness[J]. Food Chemistry, 2013,141(2): 675-679. DOI:10.3969/j.issn.1008-5467.2006.12.012.
[4] 高星, 李欣, 李铮, 等. 肌动球蛋白磷酸化对其解离的影响[J]. 食品科学, 2017, 38(9): 21-26. DOI:10.7506/spkx1002-6630-201709004.
[5] 方海田, 刘慧燕, 德力格尔桑. 宰后肌肉僵直过程中游离钙离子的作用及影响其变化的因素[J]. 肉类工业, 2007(12): 31-32.
[6] KOOHMARAIE M, GEESINK G H. Contribution of postmortem muscle biochemistry to the delivery of consistent meat quality with particular focus on the calpain system[J]. Meat Science, 2006, 74(1):34-43. DOI:10.1016/j.meatsci.2006.04.025.
[7] 张丽, 王莉, 周玉春, 等. 适宜宰后成熟时间提高牦牛肉品质[J]. 农业工程学报, 2014, 30(5): 325-331. DOI:10.3969/j.issn.1002-6819.2014.15.041.
[8] WU Gaojie, FAROUK M M, CLERENS S, et al. Effect of beef ultimate pH and large structural protein changes with aging on meat tenderness[J]. Meat Science, 2014, 98(4): 637-645. DOI:10.3969/j.issn.1002-6819.2014.15.041.
[9] 贾小翠, 李春保, 徐幸莲, 等. 禁食对僵直前后鸡肉加工特性的影响[J].食品科学, 2011, 32(19): 23-27.
[10] 王欣, 李梅, 廖志钢, 等. 尸僵再形成的实验性研究[J]. 法医学杂志,2001, 17(4): 202-204. DOI:10.3969/j.issn.1004-5619.2001.04.004.
[11] 宋洁, 侯成立, 袁有云, 等. 不同部位羊肉烤制加工适宜性研究[J].食品科学, 2017, 38(15): 108-114. DOI:10.7506/spkx1002-6630-201715018.
[12] CULLER R D, SMITH G C, CROSS H R. Relationship of myofibril fragmentation index to certain chemical, physical and sensorycharacteristics of bovine longissimus muscle[J]. Journal of Food Science, 1978, 43(4): 1177-1180. DOI:10.1111/j.1365-2621.1978.tb15263.x.
[13] KADEE J R, AVIGAIL P, WANG K K W. Casein zymography: a method to study μ-calpain, m-calpain, and their inhibitory agents[J].Archives of Biochemistry and Biophysics, 1995, 319(1): 211-216.DOI:10.1006/abbi.1995.1284.
[14] MELODY J L, LONERGAN S M, ROWE L J, et al. Early postmortem biochemical factors influence tenderness and water-holding capacity of three porcine muscles[J]. Animal Science, 2004, 82(4): 1195-1205.DOI:10.2527/2004.8241195x.
[15] 吴菊清, 李春保, 周光宏, 等. 宰后成熟过程中冷却牛肉、猪肉色泽和嫩度的变化[J]. 食品科学, 2008, 29(10): 136-139. DOI:10.3321/j.issn:1002-6630.2008.10.026.
[16] FRYLINCK L, STRYDOM P E, WEBB E C, et al. Effect of south African beef production systems on post-mortem muscle energy status and meat quality[J]. Meat Science, 2013, 93(4): 827-837.DOI:10.1016/j.meatsci.2012.11.047.
[17] 徐昶, 朱虹, 李春保, 等. 环境温度对鸡肉宰后僵直过程中ATP降解的影响[J]. 江苏农业科学, 2010(1): 257-259. DOI:10.3969/j.issn.1002-1302.2010.01.101.
[18] KOLCZAK T, POSPIECH E, PALKA K, et al. Changes in structure of psoas major and minor and semitendinosus muscles of calves, heifers and cows during post-mortem ageing[J]. Meat Science, 2003, 64(1):77-83. DOI:10.1016/S0309-1740(02)00164-X.
[19] GEESINK G, SUJANG S, KOOHMARAIE M. Tenderness of preand post rigor lamb longissimus muscle[J]. Meat Science, 2011, 88(4):723-726. DOI:10.1016/j.meatsci.2011.03.003.
[20] 李胜杰, 徐幸莲, 周光宏. 宰后肌动球蛋白解离对肉品嫩度的影响研究进展[J]. 食品科学, 2010, 31(21): 442-445.
[21] KOOHMARAIE M. Biochemical factors regulating the toughening tenderization processes of meat[J]. Meat Science, 1996, 43: 193-201.DOI:10.1016/0309-1740(96)00065-4.
[22] 刘佳东, 余群力, 李永鹏. 宰后冷却牦牛肉排酸过程中肉用品质的变化[J]. 甘肃农业大学学报, 2011, 46(2): 111-114. DOI:10.13432/j.cnki.jgsau.2011.02.002.
[23] HOPKINS D L, TOOHEY E S, LAMB T A, et al. Explaining the variation in the shear force of lamb meat using sarcomere length, the rate of rigor onset and pH[J]. Meat Science, 2011, 88(4): 794-796.DOI:10.1016/j.meatsci.2011.03.004.
[24] 师希雄, 余群力, 包富贵, 等. 牦牛肉成熟过程中钙蛋白酶对微观结构的影响[J]. 食品与机械, 2013, 29(5): 173-175.
[25] 李培迪, 李欣, 李铮, 等. 冰温贮藏对宰后肌肉成熟进程的影响[J]. 中国农业科学, 2016, 49(3): 554-562. DOI:10.3864/j.issn.0578-1752.2016.03.013.
[26] 李诚, 谢婷, 付刚, 等. 猪肉宰后冷却成熟过程中嫩度指标的相关性研究[J]. 食品科学, 2009, 30(17): 163-166. DOI:10.3321/j.issn:1002-6630.2009.17.038.
[27] 廖洪波, 明建, 贺稚非, 等. 钙蛋白酶和肉的成熟嫩化[J]. 肉类工业,2003(8): 27-29. DOI:10.3969/j.issn.1008-5467.2003.08.013.
[28] MOHRHAUSER D A, LONERGAN S M, HUFF-LONERGAN E,et al. Calpain-1 activity in bovine muscle is primarily influenced by temperature, not pH decline1[J]. American Society of Animal Science,2014, 92(3): 1261-1270. DOI:10.2527/jas2013-7270.
[29] LOMIWES D, FAROUK M M, WU G, et al. The development of meat tenderness is likely to be compartmentalised by ultimate pH[J]. Meat Science, 2014, 96(1): 646-651. DOI:10.1016/j.meatsci.2013.08.022.
[30] MARINO R, ALBENZIO M, DELLA MALVA A, et al. Changes in meat quality traits and sarcoplasmic proteins during aging in three different cattle breeds[J]. Meat Science, 2014, 98(2): 178-186.DOI:10.1016/j.meatsci.2014.05.024.
[31] KIM Y H B, LUC G, ROSENVOLD K. Pre rigor processing, ageing and freezing on tenderness and colour stability of lamb loins[J]. Meat Science, 2013, 95(2): 412-418. DOI:10.1016/j.meatsci.2013.05.017.
[32] 朱建军, 王晓宇, 胡萍, 等. 组织蛋白酶对腌肉制品风味的影响[J].食品工程, 2013(3): 4-6. DOI:10.3969/j.issn.1673-6044.2013.03.002.
Changes of Physicochemical Characteristics of Lamb Longissimus dorsi Muscle during Rigor Development and Aging
LI Guixia1,2, LI Xin2, LI Zheng2, WANG Ying1,2, ZHU Jie1,*, ZHANG Dequan2,*
(1. Laboratory of Biomechanics and Engineering, Institute of Biophysics, College of Science, Northwest A & F University,Yangling 712100, China; 2. Key Laboratory of Agro-Products Processing, Ministry of Agriculture,Institute of Food Science and Technology, Chinese Academy of Agricultural Sciences, Beijing 100193, China)
Abstract:The carcass undergoes a series of complicated physiological and biochemical processes driving the muscle to meat conversion after slaughter. Studying rigor development and postmortem aging is expected to provide theoretical supports for meat quality improvement and processing. Six crossbred rams (Small-tailed Han sheep × Beijing native sheep breed)were slaughtered, and both longissimus dorsi muscles (n = 12) from each carcass were excised and stored at 4 ℃ for 0.5,2.0, 6.0, 12.0, 24.0, 48.0, 72.0, 120.0 and 168.0 h to determine pH, ATP content, sarcomere length, shear force, myofibril fragmentation index (MFI), protein degradation and μ-calpain activity. During the conversion of muscle to meat, pH initially decreased and then remained not changed significantly at 24 h postmortem (P 〉 0.05). Shear force first increased until reaching a maximum value at 24 h after slaughter and then decreased. Sarcomere length was first shortened until reaching a minimum value at 48 h and then extended gradually. ATP content was initially increased, then decreased and finally tended to be stable. The 80 kD subunit of μ-calpain degraded almost completely disappeared at 24 h, and a 78 kD subunit was formed firstly and then degraded completely at 48 h after slaughter. Desmin and troponin T as the substrates of μ-calpain were degraded during ageing and intact proteins were scarcely be observed at 168 h. In conclusion, the development of rigor mortis in longissimus dorsi started at 2 h postmortem and reached a peak at 24 h, and it began to disappear at 48 h. The meat tenderness was improved greatly after the complete disappearance of rigor mortis.
Key words:lamb longissimus dorsi muscle; rigor; tenderness; aging
DOI:10.7506/spkx1002-6630-201721018
中图分类号:TS251.5+3
文献标志码:A
文章编号:1002-6630(2017)21-0112-07
收稿日期:2016-09-01
基金项目:国家农业科技创新工程项目;公益性行业(农业)科研专项(201303083);国家现代农业(肉羊)产业技术体系建设专项(CARS-39)
作者简介:李桂霞(1990—),女,硕士,研究方向为肉品科学与工程。E-mail:liguixia1990@163.com
*通信作者:朱杰(1980—),男,副教授,博士,研究方向为肉品科学与工程。E-mail:jiezhu@nwafu.edu.cn张德权(1972—),男,教授,博士,研究方向为肉品科学与工程。E-mail:dequan_zhang0118@126.com
引文格式:李桂霞, 李欣, 李铮, 等. 宰后僵直及成熟过程中羊背最长肌理化性质的变化[J]. 食品科学, 2017, 38(21): 112-118.
DOI:10.7506/spkx1002-6630-201721018. http://www.spkx.net.cn
LI Guixia, LI Xin, LI Zheng, et al. Changes of physicochemical characteristics of lamb longissimus dorsi muscle during rigor development and aging[J]. Food Science, 2017, 38(21): 112-118. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201721018. http://www.spkx.net.cn