表1 PCR扩增所用的引物序列
Table 1 Primer sequences used for PCR am p lifi cation
注:下划线表示酶切位点。
曾海娟,刘武康,谢曼曼,丁承超,董庆利,刘 箐*
(上海理工大学医疗器械与食品学院,上海 200093)
摘 要:在单核细胞性李斯特菌(Listeria monocytogenes)野生株EGDe actA及in lB双基因缺失株(EGDe ΔactAΔinlB)的基础上,利用同源重组的方法进一步构建了缺失营养基因dal的菌株(EGDe ΔactAΔinlBΔdal),并对该缺失菌株生长状态、毒力基因表达水平、生物被膜的形成量及细胞侵袭等方面作进一步分析。结果显示,37 ℃摇床培养6 h后,缺失株的菌浓度显著低于EGDe ΔactAΔinlB(P<0.001),培养基中补充D-丙氨酸的缺失株生长速率与亲本株相比无显著差异;实时荧光定量聚合酶链式反应结果显示,缺失株的sigB基因表达水平变化最明显(P<0.01),约下调90%;缺失株生物被膜形成量显著增加(P<0.05),培养基补充D-丙氨酸后缺失株生物被膜的生成量与亲本株相比无差异;对Coca-2细胞的侵袭无影响,表明该基因对细菌生长能力及生物被膜形成具有重要的调控作用,并不影响菌株对细胞的侵袭力。此缺失株的构建为进一步研究基因dal的功能提供了理论支持。
关键词:单核细胞性李斯特菌;基因敲除;生长能力;细胞侵袭;生物被膜
单核细胞性李斯特菌(Listeria monocytogenes)是一种重要的革兰氏染色阳性食源性致病菌,在自然环境中广泛分布[1]。单核细胞性李斯特菌具有较强的生存能力,pH 4.5~9、温度0~45 ℃均可增殖[2],在干酪、原奶、冰淇淋、生肉、海鲜及方便食品,如:熟肉、熏鱼中均有检出[3-6]。超过99%的单核细胞性李斯特菌感染是由于食用了被该菌污染的食物[7],死亡率达30%以上。单核细胞性李斯特菌是典型的胞内寄生菌,它可通过李斯特菌溶血素、磷脂酰肌醇特异性磷脂酶C和卵磷脂酶的裂解作用逃离吞噬小体[8-9],并在细胞质中增殖。单核细胞性李斯特菌可穿透肠道屏障、胎盘屏障和血脑屏障,引起哺乳动物和人的肠胃炎、败血症、孕畜流产及脑膜炎等[10]。
基因dal为营养基因,存在于多种细菌中。其编码丙氨酸消旋酶(alanine racemase,A lr)蛋白,可将L-丙氨酸转化为D-丙氨酸(D-A la),后者为细菌细胞壁肽聚糖的重要组成成分[11]。在革兰氏阳性菌中,例如芽孢杆菌属与乳杆菌属[12],仅存在一种催化生成D-A la的酶,即为A lr。而革兰氏阴性菌中,例如大肠杆菌[13],存在两种可催化产生D-A la的酶(A lr与DadX)。在单核细胞性李斯特菌中,同样存在2 种途径生成D-A la:基因dal编码A lr,可将L-A la转化为D-A la;基因dat编码D-氨基酸氨基转移酶蛋白,可将D-谷氨酸与丙酮酸通过转氨作用生成D-A la与α-酮戊二酸,当dal与dat基因全部缺失时,由于不能合成D-A la,在无外源D-A la添加时,该dal/dat双基因缺失菌株将发生溶菌死亡[14]。目前尚鲜有研究报道,当菌株缺失dal基因时,对该缺失菌株在生长状态、毒力基因表达水平、细胞侵袭性、生物被膜的生成量等方面的影响。
本研究以单核细胞性李斯特菌野生株EGDe actA及inlB双基因缺失株(EGDe ΔactAΔinlB)为亲本,利用同源重组的方法进一步构建缺失dal基因的菌株(EGDe ΔactAΔinlBΔdal),并对构建的该缺失菌株生长状态、毒力基因表达水平、细胞侵袭性及生物被膜的生成量等方面作进一步分析。该项研究对菌株EGDe ΔactAΔinlBΔdal的特性提供了依据,为研究dal基因的功能提供材料。
1.1 材料与试剂
单核细胞性李斯特菌野生株E G D e由本实验室保存,EGD e Δa c tAΔin lB由本实验室构建。大肠杆菌(Esch erich ia co li)DH 5α感受态细胞购自天根生化科技(北京)有限公司,穿梭质粒pKSV7由上海交通大学史贤明教授惠赠。
细菌总DNA提取试剂盒、胶回收试剂盒、质粒提取试剂盒 天根生化科技(北京)有限公司;实时荧光定量聚合酶链式反应(quantitative real-time polymerase chain reaction,qRT-PCR)相关试剂、PCR相关试剂、DNA连接酶、限制性内切酶 宝生物工程(大连)有限公司;脑心浸液培养基(brain heart infusion,BHI) 北京陆桥技术有限公司;氨苄青霉素、氯霉素及其他常规试剂国药集团化学试剂有限公司。
1.2 仪器与设备
PCR热循环仪、q RT-PCR仪 美国A p p lied Biosystems公司;凝胶成像仪、NANODROP浓度测定仪美国Thermo公司;M ini-power电泳仪、电穿孔仪 美国伯乐公司;SpectraM ax M 2多功能酶标仪 美国分子仪器公司。
1.3 方法
1.3.1 引物
根据GenBank登录的基因序列(Gene ID:986294),用Prim er3 Inpu t设计扩增上游同源臂引物P1/P2,下游同源臂引物P3/P4。连接后PCR引物为P1/P4,全长1 342 bp,缺失片段大小为1 202 bp。相关引物序列见表1,由生工生物工程(上海)股份有限公司合成。
表1 PCR扩增所用的引物序列
Table 1 Primer sequences used for PCR am p lifi cation
注:下划线表示酶切位点。
1.3.2 基因缺失株的构建
1.3.2.1 穿梭重组质粒的构建
利用引物P1/P2及P3/P4分别扩增dal基因的上下游同源臂片段,经Eco RⅠ单酶切后30℃进行连接16 h。以连接体系为模板,利用引物P1/P4对连接产物进行扩增。扩增产物经DNA回收试剂盒割胶纯化回收,再经SalⅠ、SmaⅠ双酶切。酶切后经割胶纯化回收,与经Sa lⅠ、SmaⅠ酶切的穿梭质粒pKSV 7连接,并转化至大肠杆菌DH5α中,涂布含氨苄青霉素(100 μg/m L)的LB固体培养基培养。
1.3.2.2 电转化与同源重组
挑取LB平板的单菌落培养,利用P1/P4进行PCR鉴定。鉴定为阳性的克隆抽提质粒后进行双酶切鉴定,并送华大基因测序。测序正确的重组质粒经11.25 kV/c m、4 ms电转化至EGDe ΔactAΔinlB中,氯霉素(10 μg/m L)抗性平板培养48 h,并用引物P1/P4对平板上的单菌落进行鉴定。鉴定为阳性的克隆在42 ℃和氯霉素双重压力下传15 代,使之发生单交换同源重组。然后在30 ℃无抗性条件下传15 代,末代培养物划线于无抗性BHI平板。
1.3.2.3 缺失株的鉴定
挑取BHI平板上的单菌落于液体培养基30 ℃培养,分别用引物P1/P4及引物dal-F/dal-R进行PCR鉴定,鉴定成功的菌株分别划线于BHI平板及含氯霉素(10 μg/m L)抗性平板,37 ℃培养。
1.3.3 缺失株生长曲线测定
为确定该基因是否会对菌株生长能力有影响,采用酶标仪对EGDe ΔactAΔinlB及敲除da l菌株EGDe ΔactAΔinlBΔdal(外源不补充及补充200 μg/m L D-A la)的生长进行测定[15-16]。挑取平板上的单菌落37 ℃过夜培养15 h后,按1∶100转接种到新鲜BHI培养基中,37℃摇床培养。每间隔1 h吸取样品于96 孔酶标板中,酶标仪测定600 nm波长处的光密度(OD600nm)后继续37 ℃摇床培养,连续测12 h。
1.3.4 毒力基因表达水平的qRT-PCR检测
表2 qRT-PCR所用的引物序列
Table 2 Primer sequences used for qRT-PCR

为确定该营养基因缺失后是否会对其他毒力基因及dat的表达量有影响,本实验选取了李斯特菌EGDe的12 个毒力基因[17]及与D-A la合成有关的2 个基因,比较了3 株菌在毒力基因表达量上存在的差异。分别挑取EGDe、EGDe ΔactAΔinlB及EGDe ΔactAΔinlBΔdal平板上的单菌落过夜培养后,各取5 m L饱和菌液提取RNA后,按照TaKaRa反转录试剂盒的操作说明先去除其中的DNA,再反转录为cDNA。以cDNA为模板进行qRTPCR,实验中选取的毒力基因引物序列参照文献[18],见表2。数据采用2-ΔΔCt法进行相对定量分析,以EGDe的基因表达水平为对照,以16S rRNA为内参。计算公式为:ΔCt=菌株目的基因Ct均值-各自内参基因Ct均值;ΔΔCt=ΔCt(敲除株及亲本株目的基因)-ΔCt(对应的EGDe目的基因)。2-ΔΔCt为毒力基因对EGDe相应毒力基因的表达量的倍数,即EGDe的每个目的基因表达水平均以1表示,EGDe ΔactAΔinlB及EGDe ΔactAΔinlBΔdal的目的基因表达水平大于1为表达上调,反之则为表达下调。
1.3.5 生物被膜生成量的测定
构成生物被膜的胞外多糖可与结晶紫结合,可利用胞外多糖的量来衡量生物被膜的生成量。操作方法根据文献报道并稍作改良后进行[19]。将过夜培养的细菌用BHI液体培养基调整至OD600nm为0.15,取200 μL菌悬液加入96 孔细胞培养板中,于37 ℃条件下培养48 h后,无菌PBS洗3 遍,150 μL 1%的结晶紫染色30 m in,无菌水洗3遍,每孔滴加170 μL 95%乙醇溶液洗脱30 m in,酶标仪测量其在570 nm波长处的吸光度。
1.3.6 细胞侵袭实验
按照相关文献报道[20],将生长良好的Caco-2细胞消化并转移至12 孔细胞培养板中,过夜培养20 h左右。细菌侵袭前,将细胞培养板中完全培养基吸出,换成不含血清的不完全培养基,然后按细菌和细胞100∶1的比例分别加入新鲜的野生株EGDe、亲本株EGDe ΔactAΔinlB及敲除株EGDe ΔactAΔinlBΔdal共培养3 h。侵袭结束后,换用庆大霉素(200 μg/m L)37 ℃处理30 m in。将庆大霉素吸出,无菌生理盐水轻柔清洗2 次,再用1% Triton X-100裂解细胞,释放细菌。每孔充分吹打并收集至离心管中,生理盐水梯度稀释涂布BHI平板,置于37 ℃培养箱培养并计数。
2.1 上下游片段扩增及连接
上下游同源臂的PCR扩增产物经1.5%琼脂糖凝胶电泳,条带大小分别为607 bp和735 bp左右(图1a),连接后PCR扩增产物经电泳,显示一条约1 342 bp的条带(图1b),与预期结果一致。
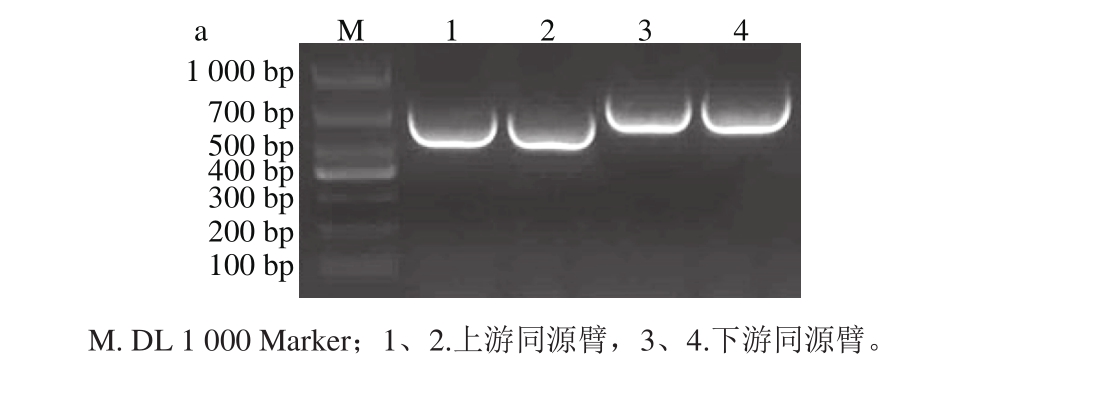

图1 上下游同源臂(a)及连接后片段(b)大小
Fig. 1 Fragment sizes of separate and connected upstream and downstream homologous arm s
2.2 穿梭重组载体构建及双酶切鉴定结果
连接后片段经SalⅠ、SmaⅠ双酶切后,与同样酶切的穿梭质粒pKSV7连接,并转化至大肠杆菌DH5α中。阳性克隆测序成功后,经双酶切鉴定,片段大小与预期一致(图2a),表明重组载体构建成功。抽提质粒电转化至EGDe ΔactAΔinlB中,引物P1/P4进行PCR鉴定,可见1 342 bp左右的条带(图2b泳道1~3),表明重组质粒已经电转化至EGDe ΔactAΔinlB中。
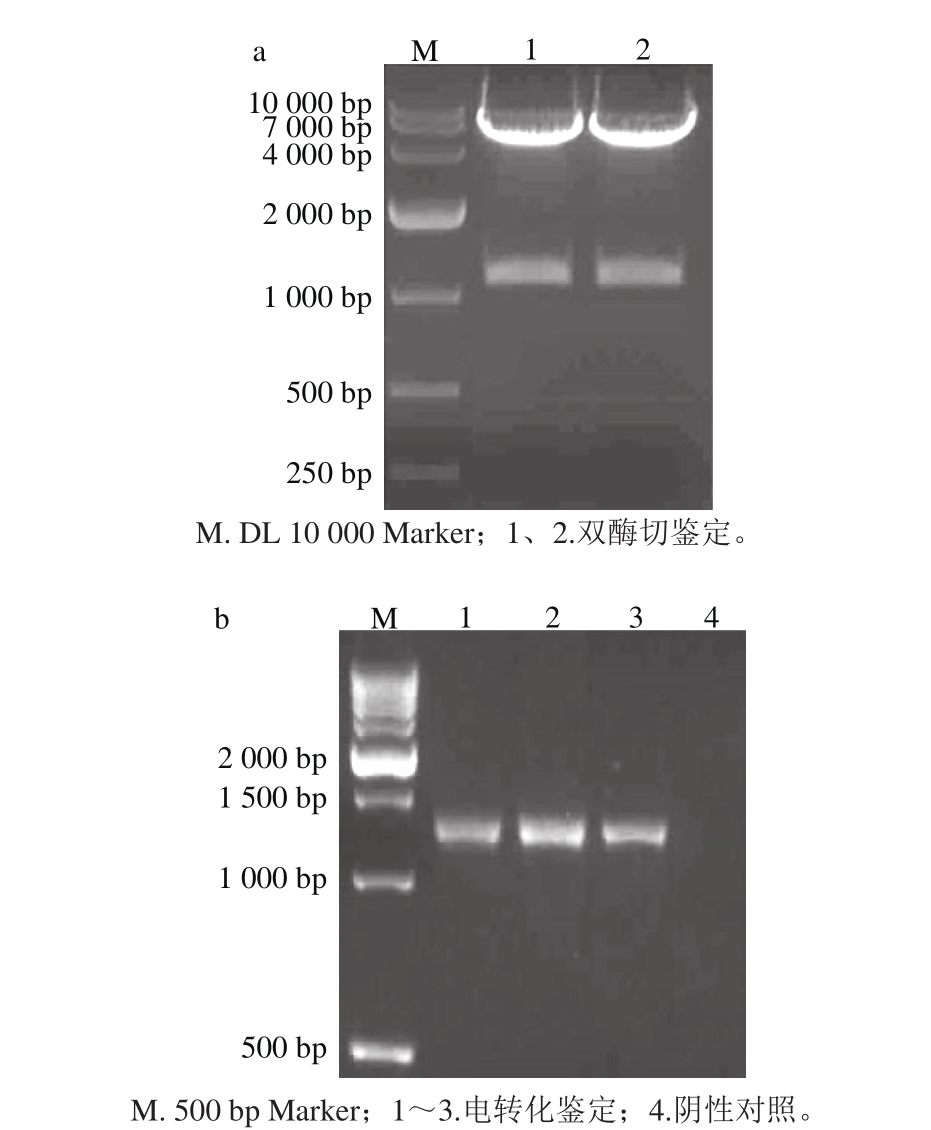
图2 双酶切(a)及电转化(b)鉴定结果
Fig. 2 Identifi cation by doub le enzyme digestion and electrotransform ation
2.3 缺失株的鉴定
dal-F/dal-R的结合位点位于上游同源臂引物P2及下游同源臂引物P3之间1 202 bp的序列上,若dal基因被敲除,则该引物将不会扩增出条带。鉴定时,采用引物P1/P4及引物dal-F/dal-R对单菌落分别进行PCR扩增,若P1/P4扩增出1 342 bp左右的条带,而dal-F/dal-R未扩增出条带,则该菌株为疑似敲除菌株。如图3a所示,引物P1/P4扩增出1 342 bp左右的条带(泳道3),而未敲除菌株(泳道1、2)扩增出约2 544 bp的条带;如图3b所示,引物dal-F/dal-R未扩增出条带(泳道3),而未敲除株扩增出约428 bp的条带(泳道1、2),表明3号为疑似敲除株,经氯霉素抗性进一步鉴定,该克隆不能在抗性平板上生长,确定3号为敲除株。
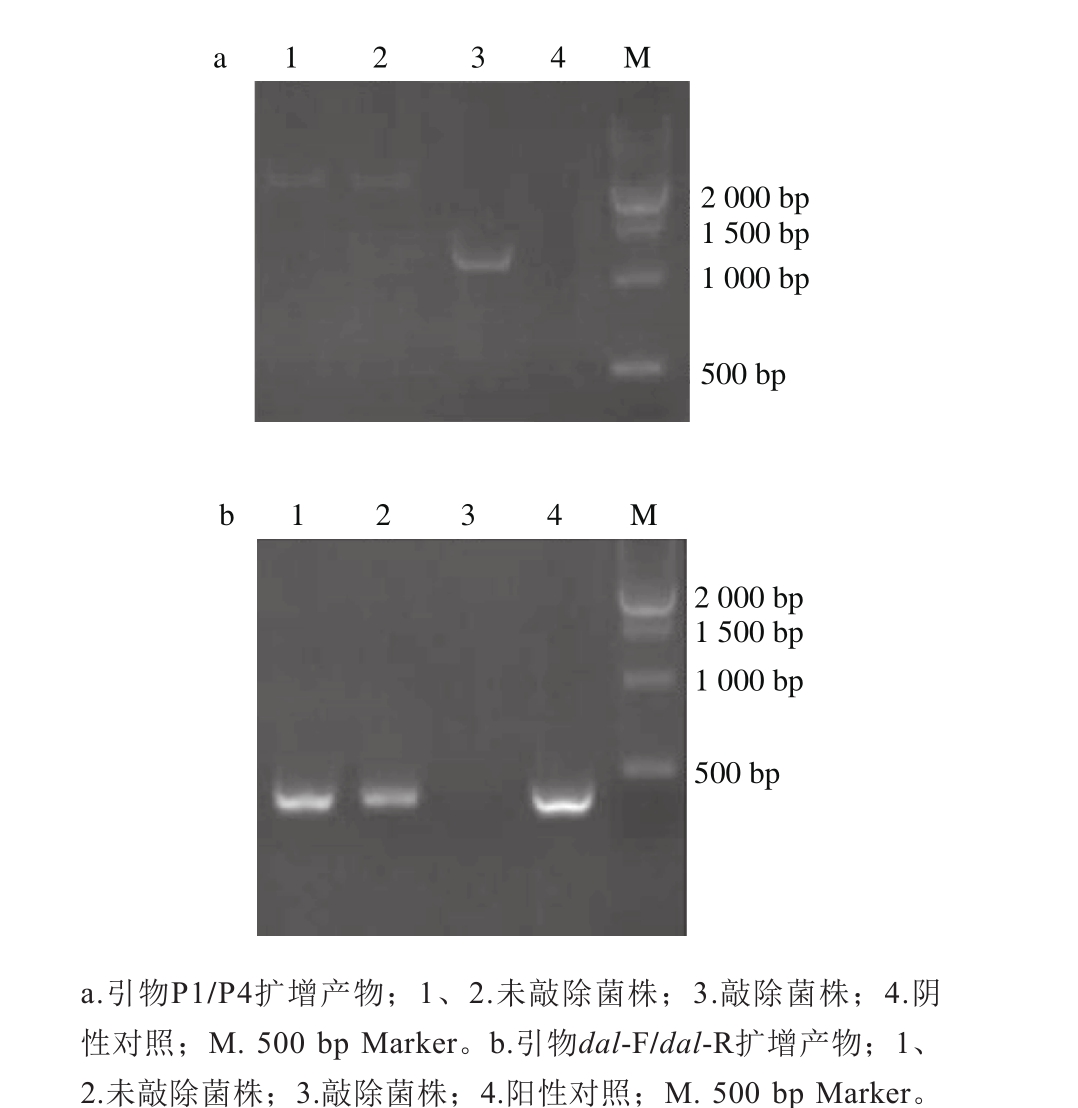
图3 基因缺失株的鉴定
Fig. 3 Identification of dal deletion mutant
2.4 生长曲线分析

图4 生长曲线
Fig. 4 Grow th curves
酶标仪测定连续12 个时间点的OD600nm值绘制生长曲线,如图4所示,与亲本株EGDe ΔactAΔinlB相比,敲除株EGDe ΔactAΔinlBΔdal从培养6 h的对数生长期到12 h平台期达到的菌浓度显著降低(P<0.001),而在敲除株培养基中添加D-A la后生长趋势与亲本株相比无差别。表明尽管李斯特菌可通过另一途径经丙氨酸转氨酶作用产生D-Ala,但dal基因的缺失使菌株的生长能力降低。
2.5 毒力基因表达结果
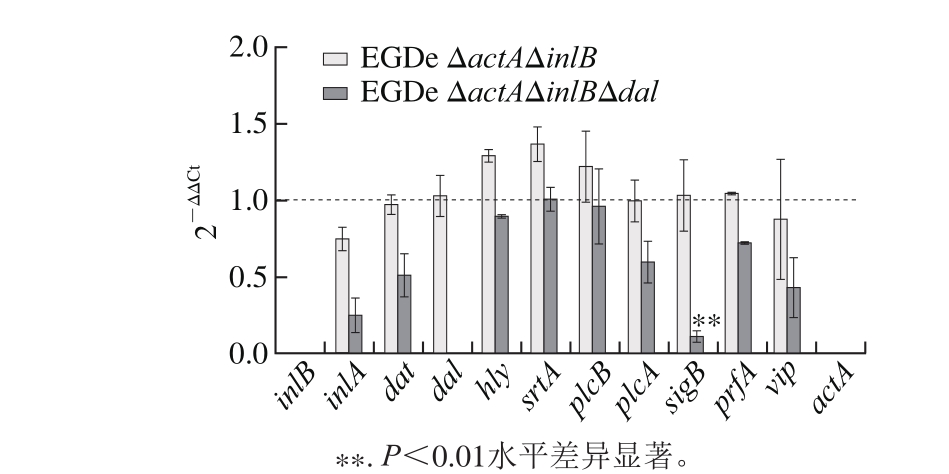
图5 qRT-PCR结果
Fig. 5 Viru lence gene exp ression levels detected by RT-PCR
由图5可知,菌株EGDe ΔactAΔinlBΔdal的dal基因无信号,进一步验证dal基因的缺失;EGDe ΔactAΔinlB与EGD e相比,未有显著性差异(P>0.0 5);EGDe ΔactAΔinlBΔdal与EGDe ΔactAΔinlB相比,基因表达量均略微下调,其中sigB基因表达水平变化最明显(P<0.01),约下调90%。
2.6 生物被膜生成量分析
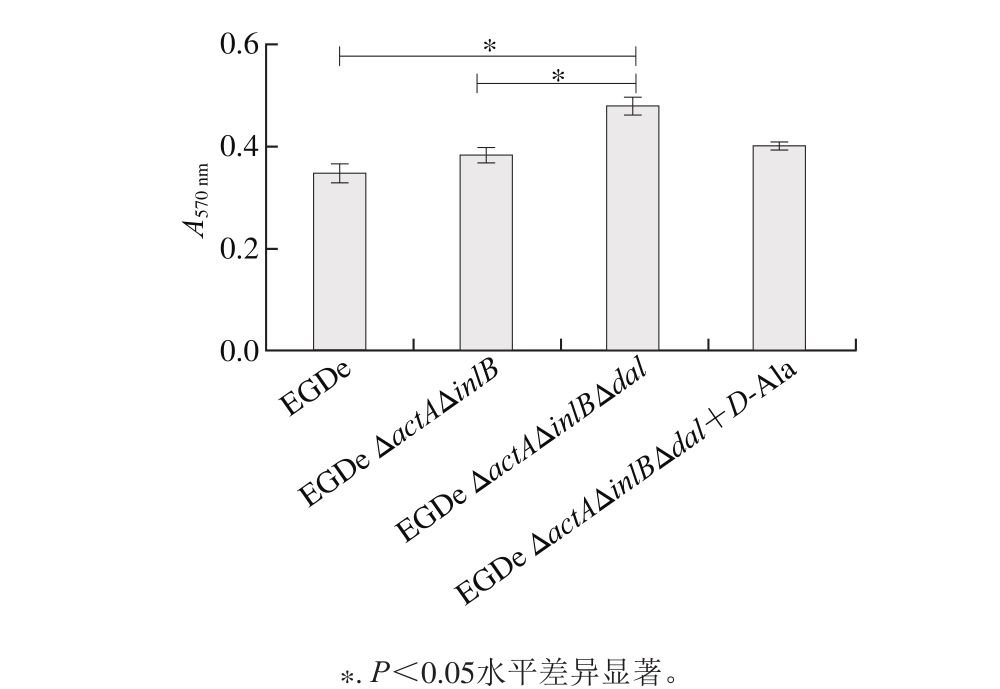
图6 生物被膜生成量比较
Fig. 6 Comparison of biofi lm formation between the wild-type and the mutant strains
生物被膜是细菌抵抗外界不利环境时形成的,它是一种多细胞聚合物,由细菌团块与表多糖、蛋白和胞外DNA构成的细胞外基质形成的复合物。由图6可知,dal缺失株与野生株EGDe及亲本株EGDe ΔactAΔinlB相比,形成生物被膜的能力显著增加(P<0.05),而在培养基中添加D-A la后缺失株菌膜生成量与EGDe及EGDe ΔactAΔinlB相比均无显著差异,推测由于dal基因的缺失使菌株处于D-A la营养不足的胁迫状态,因而对于外界不利条件形成自我保护的生物被膜。
2.7 细胞侵袭结果
由图7敲除株对Caco-2细胞侵袭结果可以看出,与野生株相比敲除actA及inlB基因后,菌株EGDe ΔactAΔinlB的侵袭能力显著下降(P<0.01),表明actA及inlB基因在细胞侵袭过程中发挥了重要作用[21]。但dal基因敲除后,与菌株EGDe ΔactAΔinlB相比,并未对侵袭结果产生显著影响,表明dal基因敲除后,菌株对细胞的侵袭力并无发生变化,也不会引起菌株毒力返祖的现象。
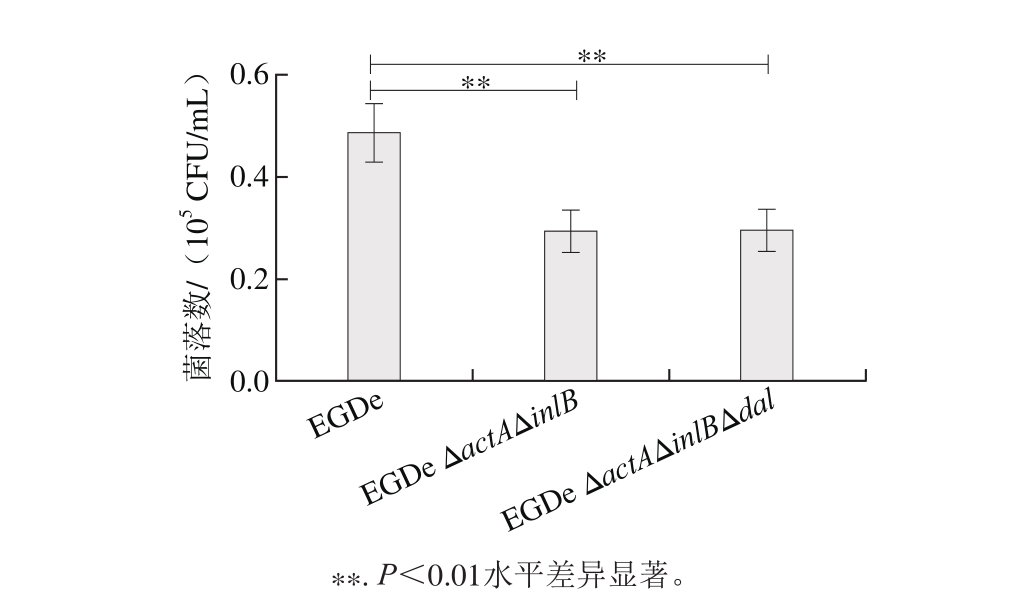
图7 细胞侵袭结果
Fig. 7 Com parison of cell invasion ability between the w ild-type and the mutant strains
单核细胞性李斯特菌识别、黏附与侵入宿主细胞的过程与内化素基因(inl)分泌的内化素A和B有关[22-23],而其细胞内增殖及细胞间扩散能力则由毒力基因plcA、plcB、hly、prfA、actA等介导[24-26]。本研究在单核细胞性李斯特菌actA及inlB双基因缺失株的基础上成功构建了dal基因缺失的三基因缺失株,并对该缺失菌株的生长特性、毒力基因表达的变化、生物被膜的生成量及对细胞的侵袭性进行了研究。结果显示,缺失dal基因的菌株在无外源D-A la补充时,虽不影响菌株的生存,但从生长曲线可以看出,缺失株6 h后菌浓度低于亲本株,而外源补充D-A la后生长能力与亲本株相比无差异,表明通过丙氨酸转氨作用单一途径合成D-A la不足以维持菌株原有的生长繁殖速率,使得菌浓度降低。qRT-PCR结果显示,sigB基因表达水平约下调90%。由于sigB基因在许多革兰氏阳性菌,例如芽孢杆菌属、李斯特菌和金黄色葡萄球菌中,作为应激反应的中心调节子[27-28],推断菌株EGDe ΔactAΔinlBΔdal在外界不利条件下生长能力可能有一定影响。缺失株生物被膜生成量显著增加(P<0.05),但在培养基中添加D-A la后生物被膜生成量与亲本株相比无差异,表明基因dal缺失使菌株处于一种外界营养胁迫状态,该胁迫对菌体生长是不利的,因而产生了应对不利环境的菌膜。该缺失株的构建表明dal基因在菌株生长及生物被膜形成上起重要的调控作用,但不会对菌株毒力产生影响。
在革兰氏阳性菌如鼠伤寒沙门菌、猪霍乱沙门菌中[29-30],同样存在类似功能的基因asd,编码天冬氨酸β-半乳糖脱氢酶。asd缺失菌株在无外源二氨基庚二酸存在的条件下会发生溶菌死亡,通过质粒回补asd基因并携带外源抗原基因而表达相应抗原蛋白,能有效解决质粒携带外源基因表达不稳定的问题,为沙门菌活疫苗载体的研发开辟了新的途径。本研究的下一步实验是进一步缺失dat基因,并通过质粒回补dal基因并携带外源抗原基因,构建能稳定表达外源基因的单核细胞性李斯特菌活疫苗载体。研究通过对缺失株生长曲线、qRT-PCR、生物被膜的生成量、细胞侵袭实验的研究,为基因dal缺失后菌株的生物学特性提供了依据,对进一步缺失dat基因,构建李斯特菌活疫苗载体提供了理论支持。
参考文献:
[1] REDA W W, ABDEL-MOEIN K, HEGAZI A, et al. Listeria monocytogenes: an emerging food-borne pathogen and its public health implications[J]. Journal of Infection in Developing Countries,2016, 10(2): 149-154. DOI:10.3855/jidc.6616.
[2] HA IN T, CHATTERJEE S S, GHA I R, et al. Pathogenom ics of Listeria spp.[J]. International Journal of Medical M icrobiology, 2007,297: 541-557. DOI:10.1016/j.ijmm.2007.03.016.
[3] QUERO G M, SANTOVITO E, V ISCONTI A, et al. Quantitative detection of Listeria monocytogenes, in raw m ilk and soft cheeses:culture-independent versus liquid- and solid-based culture-dependent real time PCR approaches[J]. Food Science & Technology Research,2014, 58(1): 11-20. DOI:10.1016/j.lw t.2014.03.005.
[4] MASTRONICOLIS S K, DIAKOGIANNIS I, BERBERI A, et al.Eff ect of cold adaptation on the survival of Listeria monocytogenes,in ice-cream formulations during long-term frozen storage[J]. Annals of Microbiology, 2011, 61(4): 931-937. DOI:10.1007/s13213-011-0216-7.
[5] PESAVENTO G, DUCCI B, N IERI D, et al. Prevalence and antibiotic susceptibility of Listeria spp. isolated from raw meat and retail foods[J]. Food Control, 2010, 21(5): 708-713. DOI:10.1016/j.foodcont.2009.10.012.
[6] GUSMAN V, MEDIC D, JELESIC Z, et al. Listeria monocytogenes isolated in ready-to-eat food in south Backa region of Vojvodina province, Serbia[J]. Archives of Biological Sciences, 2014, 66(1): 11-14. DOI:10.2298/ABS1401011G.
[7] ALLEN K J, WALECKA-ZACHARSKA E, CHEN J C, et al. Listeria monocytogenes-an exam ination of food chain factors potentially contributing to antim icrobial resistance[J]. Food M icrobiology, 2015,54: 178-189. DOI:10.1016/j.fm.2014.08.006.
[8] LAM G, FATTOUH R, MUISE A, et al. Listeriolysin O suppresses phospholipase C-mediated activation of the m icrobicidal NADPH oxidase to promote Listeria monocytogenes infection[J]. Cell Host &M icrobe, 2011, 10(6): 627-634. DOI:10.1016/j.chom.2011.11.005.
[9] YEUNG P S, ZAGORSKI N, MARQUIS H. The metalloprotease of Listeria monocytogenes controls cell wall translocation of the broadrange phospholipase C[J]. Journal of Bacteriology, 2005, 187(8):2601-2608. DOI:10.1128/JB.187.8.2601-2608.2005.
[10] LOW J C, DONACHIE W. A review of Listeria monocytogenes, and listeriosis[J]. Veterinary Journal, 1997, 153(1): 9-29. DOI:10.1016/S1090-0233(97)80005-6.
[11] AWASTHY D, BHARATH S, SUBBULAKSHM I V, et al. Alanine racemase mutants of Mycobacterium tuberculosis require D-alanine for grow th and are defective for survival in macrophages and m ice[J].M icrobiology, 2012, 158(2): 319-327. DOI:10.1099/mic.0.054064-0.
[12] STEEN A, PALUMBO E M, COCCONCELLI P S, et al. Autolysis of Lactococcus lactis is increased upon D-alanine dep letion of peptidoglycan and lipoteichoic acids[J]. Journal of Bacteriology, 2005,187(1): 114-124. DOI:10.1128/JB.187.1.114-124.2005.
[13] W ILD J, HENNIG J, LOBOCKA M, et al. Identifi cation of the dadX,gene coding for the predom inant isozyme of alanine racemase in Escherichia coli K12[J]. MGG-Molecular and General Genetics, 1985,198(2): 315-322. DOI:10.1007/BF00383013.
[14] ZHAO X, LI Z, GU B, et al. Pathogenicity and immunogenicity of a vaccine strain of Listeria monocytogenes that relies on a suicide plasm id to supply an essential gene product[J]. Infection & Immunity,2005, 73(9): 5789-5798. DOI:10.1128/IAI.73.9.5789-5798.2005.
[15] YANG Y, HOU J, LIN Z, et al. Attenuated Listeria monocytogenes as a cancer vaccine vector for the delivery of CD24, a biomarker for hepatic cancer stem cells[J]. Cellular and Molecular Immunology,2014, 11(2): 184. DOI:10.1038/cm i.2013.64.
[16] 康美琴, 蔡雪薛, 谈卫军, 等. 单核细胞增生李斯特菌hfq基因缺失株的构建及其生物学特性[J]. 微生物学报, 2015, 55(4): 433-439.DOI:10.13343/j.cnki.wsxb.20140342.
[17] 亢春雨, 于宏伟, 郭润芳, 等. 单增李斯特氏菌Lm319主要毒力基因的时序性表达研究[J]. 农业生物技术学报, 2015, 23(6): 788-797.DOI:10.3969/j.issn.1674-7968.2015.06.011.
[18] 刘武康, 陈国薇, 吴嫚, 等. inlA和inlB基因缺失对单核细胞增生性李斯特菌侵袭HT29结肠癌细胞的影响[J]. 食品科学, 2016, 37(23):166-172. DOI:10.7506/spkx1002-6630-201623028.
[19] DJORDJEVIC D, W IEDMANN M, MCLAMDSBOROUGH L A.M icrotiter p late assay for assessment of Listeria monocytogenes biofi lm formation[J]. Applied & Environmental M icrobiology, 2002,68(6): 2950. DOI:10.1128/AEM.68.6.2950-2958.2002.
[20] YAMADA F, UEDA F, OCH IA I Y, et al. Invasion assay o f Listeria monocytogenes, using Vero and Caco-2 cells[J]. Journal of M icrobiological Methods, 2006, 66(1): 96-103. DOI:10.1016/j.m imet.2005.10.017.
[21] 丁承超, 曾海娟, 钟菲菲, 等. 双基因敲除减毒单增李斯特菌(ΔactA/ΔinlB)的构建及其生物学初步鉴定[J]. 现代食品科技, 2016(7): 66-71.DOI:10.13982/j.m fst.1673-9078.2016.7.011.
[22] SCHAIK W V, ABEE T. The role of σ B, in the stress response of Gram-positive bacteria-targets for food preservation and safety[J].Current Opinion in Biotechnology, 2005, 16(2): 218-224. DOI:10.1016/j.copbio.2005.01.008.
[23] GRUNDLER T, QUEDNAU N, STUMP C, et al. The surface proteins InlA and InlB are interdependently required for polar basolateral invasion by Listeria monocytogenes in a human model of the bloodcerebrospinal fluid barrier[J]. M icrobes and Infection, 2013, 15(4):291-301. DOI:10.1016/j.m icinf.2012.12.005.
[24] CARRERO J A, CALDERON B, UNANUE E R. Listeriolysin O from Listeria monocytogenes is a lymphocyte apoptogenic molecule[J].Journal of Immunology, 2004, 172(8): 4866-4874. DOI:10.4049/jimmunol.172.8.4866.
[25] HERAS A D L, CAIN R J, BIELECKA M K, et al. Regulation of Listeria virulence: PrfA master and commander[J]. Current Opinion in M icrobiology, 2011, 14(2): 118-127. DOI:10.1016/j.m ib.2011.01.005.
[26] LE MONNIER A, AUTRET N, JOIN-LAMBERT O F, et al. ActA is required for crossing of the fetop lacental barrier by Listeria monocytogenes[J]. Infection & Immunity, 2007, 75(2): 950-957.DOI:10.1128/IAI.01570-06.
[27] SOMOLINOS M, ESPINA L, PAGAN R, et al. sigB absence decreased Listeria monocytogenes EGD-e heat resistance but not its pulsed electric fields resistance[J]. International Journal of Food Microbiology,2010, 141(1/2): 32-38. DOI:10.1016/j.ijfoodm icro.2010.04.023.
[28] GAO X, LORINCZI M, HILL K S, et al. M et receptor tyrosine kinase degradation is altered in response to the leucine-rich repeat of the Listeria invasion protein internalin B[J]. Journal of Biological Chem istry, 2009, 284(2): 774-783. DOI:10.1074/jbc.M 805989200.
[29] 李静, 陈松彪, 余祖华, 等. 鼠伤寒沙门菌SL1344株环化腺苷酸合成酶缺失株平衡致死系统的构建及其雏鸡免疫保护试验[J]. 微生物学报, 2015, 55(7): 942-948. DOI:10.13343/j.cnki.wsxb.20140559.
[30] 赵战勤, 徐引弟, 吴斌, 等. 猪霍乱沙门氏菌∆asd C500株的生物学特性及作为活疫苗表达载体的应用[J]. 生物工程学报, 2009, 25(1):29-36. DOI:10.3321/j.issn: 1000-3061.2009.01.005.
Knockout of dal Gene and Its Effect on Listeria monocytogenes
ZENG Haijuan, LIU Wukang, XIE Manman, DING Chengchao, DONG Qingli, LIU Qing*
(School of Medical Instrument and Food Engineering, University of Shanghai for Science and Technology, Shanghai 200093, China)
Abstract:In this study, the actA and inlB double gene deletion mutant (EGDe ΔactAΔinlB) of Listeria monocytogenes w ild-type (WT) strain EGDe was used as the parent to delete the dal gene by homologous recombination technology, and the biological characteristics of the dal-deficient mutant such as grow th capacity, virulence gene expression, cell invasion and biofilm formation were further studied. Grow th curves showed that the concentration of the new mutant strain was significantly lower than that of EGDe ΔactAΔinlB after 6 h of shaking culture at 37 ℃ (P < 0.001), but there was no diff erence in the grow th rates of the parental and the mutant strains when D-alanine was added to the medium. Quantitative real-time-PCR showed that the sigB gene expression level of the mutant strain was changed most significantly (P < 0.01) and down-regulated by 90% compared w ith EGDe ΔactAΔinlB. The biofi lm formation of the mutant strain increased compared w ith EGDe ΔactAΔinlB, but this diff erence did not exist when D-alanine was added to the medium for the mutant. There was no significant diff erence in Caco-2 cells invasion ability compared w ith EGDe ΔactAΔinlB. The results indicated that the dal gene played an important regulatory role in the grow th and biofi lm formation of bacteria and did not aff ect the ability of cell invasion. The construction of this deletion strain can provide a tool for further study of the function of the dal gene.
Key words:Listeria monocytogenes; gene knockout; grow th capacity; biofi lm; cell invasion
DOI:10.7506/spkx1002-6630-201722008
中图分类号:TS201.3
文献标志码:A
文章编号:1002-6630(2017)22-0048-06
引文格式:
曾海娟, 刘武康, 谢曼曼, 等. 单核细胞性李斯特菌基因dal的敲除及其生物学特性初步分析[J]. 食品科学, 2017, 38(22):
48-53.
DOI:10.7506/spkx1002-6630-201722008. http://www.spkx.net.cn
ZENG Haijuan, LIU Wukang, XIE Manman, et al. Knockout of dal gene and its effect on Listeria monocytogenes[J]. Food
Science, 2017, 38(22): 48-53. (in Chinese w ith English abstract) DOI:10.7506/spkx1002-6630-201722008. http://www.spkx.net.cn
收稿日期:2016-12-29
基金项目:国家自然科学基金面上项目(31371776)
作者简介:曾海娟(1990—),女,博士研究生,研究方向为食源性致病菌致病机理。E-mail:zenghaijuan12@126.com
*通信作者:刘箐(1970—),男,教授,博士,研究方向为食源性致病菌致病机理及快速检测技术。E-mail:liuq@usst.edu.cn