朱力杰,陈 宁,陈妍婕,刘秀英,王 勃,刘 贺,何余堂,马 涛*
(渤海大学食品科学与工程学院,生鲜农产品贮藏加工及安全控制技术国家地方联合工程研究中心,辽宁 锦州 12 1013)
摘 要:将植物乳杆菌、鼠李 糖乳杆菌、嗜酸乳杆菌、干酪乳杆菌4 种乳酸菌经三三组合后作为复合发酵剂对发芽糙米乳进行发酵,通过分析酸度、活菌数、脱水收缩作用敏感性、蛋白质分解力、流变学性质等实验结果,对各乳酸菌组合的发酵特性进行评价,最终筛选出适合发酵发芽糙米乳的最佳菌株组合。结果表明:植物乳杆菌-嗜酸乳杆菌-干酪乳杆菌组合所得到的发酵发芽糙米乳具有较 高的品质。该发酵糙米乳在4 ℃条件下贮藏21 d后酸化程度较弱,下降了0.71 个单位;活菌数变化小且冷藏期间数量一直高于8.7(lg(CFU/mL)),游离氨基酸平均含量达0.86 mmol/L,流变学性质显示其剪切稀化作用较弱。故该乳酸菌组合较为适合发酵发芽糙米乳。
关键词:乳酸菌;混合发酵;发芽糙米乳;特性
随着精白米、精面粉等消费的普及,其营养物质组成单一、人体必需微量元素含量低下等缺陷逐渐引起了广大消费者的重视,因此对粗杂粮食品的追求成为了近年来主食消费的趋势之一。发芽糙米是将稻谷去掉外壳后,经浸泡、发芽,得到芽长0.5~1 mm,由幼芽、糠层和胚乳组成的糙米制品[1-2]。糙米在发芽的过程中,原有的部分营养成分含量发生变化,其中维生素、矿物质、膳食纤维的含量显著增加,糙米多酚等生理活性物质含量明显增加[3],特别是γ-氨基丁酸的含量从发芽前的0.82~5.84 mg/100 g跃升至发芽后的22.68~88.36 mg/100 g[4-5]。此外,淀粉等储能物质在发芽的过程中发生代谢消耗,因此发芽糙米及其制品的热量较发芽前有所减少。发芽改变了稻谷中部分原有组分的存在状态,使其更易消化和吸收。研究表明,发芽糙米蛋白具有一定的抗氧化活性[6],常食发芽糙米有利于预防头痛、结肠癌、心脏病,可以起到降血压、缓解便秘、调节血糖水平等作用[7-10]。基于此,食品从业者以发芽糙米为基质,研发了发芽糙米黑豆复合饮料、速食发芽糙米、发芽糙米面包、发芽糙米煎饼等多种新型糙米食品[11-14]。
为了更好地推广发芽糙米制品,丰富相关食品品种,本研究以发芽糙米粉为基质,采用复合乳酸菌(lactic acid bacteria,LAB)作为发酵剂制成发芽糙米发酵乳。该产品综合了粗粮与乳酸菌各自的优点,是一种高蛋白、高膳食纤维、高活菌数的植物发酵乳品,既保留了发芽糙米中的主要营养物质,又改进了适口性和滋气味。前期研究比较了不同乳酸菌在发芽糙米乳中发酵特性的异同[15],但使用单一菌种制成的发酵糙米乳往往不能实现品质稳定与优良风味的兼顾。陈海旭等[16]的研究发现,多菌复配发酵制成的糙米饮料在酸度、活菌数等各项指标的表现上更加均衡。因此本研究从实际生产角度出发,将不同类型的乳酸菌菌株进行三三组合,通过探究复合乳酸菌在发芽糙米乳中的发酵特性,筛选出性能优良、适合推广应用的菌株组合。
1.1 材料与试剂
发芽糙米粉、大豆分离蛋白 本溪寨香生态农业有限公司;植物乳杆菌(Lactobacillus plantarum,Lp)、鼠李糖乳杆菌(Lactobacillus rhamnosus,Lr)、嗜酸乳杆菌(Lactobacillus acidophilus,La)和干酪乳杆菌(Lactobacillus casei,Lc) 四川高福记生物科技有限公司;MRS培养基 北京中科质检生物技术有限公司;其他试剂均为分析纯 国药集团化学试剂有限公司。
1.2 仪器与设备
PHS-3C酸度计 上海雷磁仪器厂;UV-2550紫外-可见分光光度计 杭州英斯特科技有限公司;SIS60-70均质机 上海申鹿均质机有限公司;Czone5F抑菌圈测量及菌落计数仪 杭州讯数科技有限公司;DHR-1流变仪 美国TA公司。
1.3 方法
1.3.1 菌种的活化与母发酵剂的制备
分别将4 种乳酸菌的菌粉按1∶1 000(g/mL)的比例接种于三角瓶内的液体MRS培养基中,于37 ℃条件下恒温培养12 h;再以体积分数3%接种于试管内的液体MRS培养基中,37 ℃条件下恒温培养12 h,经过2 次活化后,保证活菌数至9(lg(CFU/mL))以上。随后按体积分数1%将各菌株经三三组合后接种于无菌发芽糙米乳中,37 ℃恒温培养5 h,凝乳后即为母发酵剂,置于4 ℃冰箱中保存备用。
1.3.2 技术路线
发芽糙米粉→中温α-淀粉酶酶解(质量分数0.05%,65 ℃,40 min)→煮沸(100 ℃,10 min,终止酶解)→过滤去除废渣(200 目滤布)→添加大豆分离蛋白→均质(20 MPa,10 min)→蔗糖调配(70 g/L)→灭菌(90 ℃,10 min)→冷却室温→接种母发酵剂(体积分数5%)→发酵(37 ℃,24 h)→冷藏后熟(4 ℃,12 h)→成品发芽糙米乳
在发酵乳制成后的第0、1、7、14、21天取样检测相关各项指标。
1.3.3 pH值测定
取出于4 ℃冷藏的发酵糙米乳样品置于室温条件下,待温度恢复至室温后用玻璃棒搅拌均匀,由酸度计直接测定pH值。
1.3.4 滴定酸度测定
参照GB 5009.239—2016《食品酸度的测定》方法[17],称取10.000 g搅拌均匀的发酵糙米乳于150 mL锥形瓶中,加入20 mL新煮沸冷却至室温的蒸馏水和2 mL 0.5%酚酞乙醇指示液于试样中,混合均匀后用0.1 mol/L氢氧化钠标准溶液滴定至微红色,且30 s内不褪色。记下NaOH标准溶液消耗的体积,通过计算得滴定酸度(吉尔涅尔度,ºT)。
1.3.5 活菌数测定
采用无菌生理盐水对发酵糙米乳进行梯度稀释,选取3 个合适梯度,采用MRS培养基平板倾注,于37 ℃条件下培养48 h进行活菌计数。
1.3.6 脱水收缩敏感性(susceptibility to syneresis,STS)测定
称量发酵糙米乳样品20.0 g,置于带有滤纸的漏斗中,4 ℃条件下放置2 h,收集滤液,称质量[18]。STS按公式(1)计算。
1.3.7 蛋白质分解力测定
蛋白质分解力的测定采用邻苯二甲醛比色法[19-20],在340 nm波长处测定发酵糙米乳样品的吸光度,确定蛋白质分解力。
1.3.8 流变性分析
发酵糙米乳搅拌均匀后,选用直径40 mm的不锈钢平板探头,平板与底面的间隙设置为0.5 mm,测试温度控制在25 ℃进行测定。剪切速率从0.01 s-1增大到150 s-1,在150 s-1保持120 s,然后从150 s-1减小至0.01 s-1,对得到的表观黏度、贮藏模量、损失模量、稠度系数、流动行为指数等指标进行分析。
以改进的Ostwald-de Waele模型分析发酵糙米乳下行线的流变参数,模型方程如公式(2)。
式中:η为黏度/(Pa·s);γ为剪切速率/s-1;K为稠度系数/(Pa·sn);n为流动行为指数。
2.1 pH值测定结果

图1 复合乳酸菌发酵糙米乳pH值变化
Fig. 1 Changes in pH of brown rice milk during fermentation by mixed LAB
由于目前欠缺对发酵糙米乳中各项指标进行检测的国家标准,故本实验部分沿用了国家标准在发酵牛乳中检测相关指标的方法,实 测效果较好,能够反映出发酵糙米乳在各个时期中发生的品质变化。pH值是评价发酵糙米乳品质的一项重要指标,对产品的口感和品质都会产生一定的影响,其测定简便易行,被广泛用于监测发酵乳的酸度。乳酸菌在发酵糙米乳的过程中会产生乳酸等酸性物质,致使pH值不断下降,酸度不断上升。在发酵24 h后进入冷藏后熟阶段,经观察发现前7 d乳酸菌增殖较快,酸性物质的增加速度也较快,pH值下降较多。而在后期随着发酵糙米乳体系酸度逐渐升 高,乳酸菌的生长繁殖受到抑制,因而酸性物质的产生速率放缓,限制了pH值继续快速下降,使得pH值逐渐趋于稳定。孙懿琳[21]以保加利亚乳杆菌发酵的酸乳研究结果表明前4 d是菌株产酸的关键时期,这与本研究结果一致。此外菌株之间存在协同关系,产酸速率较前期单菌实验有所增加[15],4 ℃贮藏至21 d时,pH值下降到3.82。其中Lp-La-Lc为贮藏过程中乳酸菌继续发酵产生后酸化现象最弱的组合,pH值下降了0.71。
2.2 滴定酸度变化
采用3 种复合乳酸菌发酵可以很好地弥补部分菌株产酸不足的缺陷,并使产品的滴定酸度在发酵结束时仍然处于较低水平。与单菌种发酵糙米乳相比,三菌种复合乳酸菌作为发酵剂在发酵结束的滴定酸度同样维持在较低水平[15],可见复合菌种发酵并没有提高发酵糙米乳的酸度,有研究表明这与发酵所用菌种、原料乳的种类以及发酵温度有关[22]。在前7 d中,Lp-Lr-La和Lp-La-Lc组合的酸度变化情况相近且增幅较大,而Lp-Lr-Lc和Lr-La-Lc组合的酸度变化情况接近但增幅较小;7 d以后酸度增加趋势减缓,后酸化程度减弱。其中Lr-La-Lc后酸化程度最严重,21 d内增加了38.28 ºT,另外3 种差异不大,Lp-La-Lc酸度增加最小为29.62 ºT。这与pH值的变化一致,二者显著相关(P<0.05)。
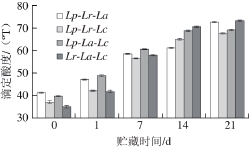
图2 复合乳酸菌发酵糙米乳滴定酸度变化
Fig. 2 Change in titratable acidity during fermentation of brown rice milk by mixed LAB
2.3 活菌数变化
表1 复合乳酸菌活菌数变化
Table 1 Changes in viable cell count during storage of fermented brown rice milk(lg(CFU/mL))

整个贮藏期间各组的乳酸菌活菌数均较高,超过了8.5(lg(CFU/mL))。乳酸菌的生长趋势为先上升后下降,基本在前7 d活菌数较高,达到最高值9(lg(CFU/mL))。贮藏前期,虽然低温条件下乳酸菌仍然可以缓慢生长,但是可能因为发酵糙米乳前期酸度增幅较大,代谢产物的积累等造成后期乳酸菌活菌数下降[23],致使其pH值、滴定酸度变化较小。欧盟规定了益生菌产品在货架期内活菌数至少要保持在6(lg(CFU/mL))以上[24],而本研究制成的复合乳酸菌发酵糙米乳中活菌数高于这一数值100 倍以上,完全符合规定。不同菌种组合在冷藏期内的活菌数存在差异,其中Lp-Lr-Lc和Lp-La-Lc组合的活菌数相对较高,可能与这两组中Lp和Lc 2种乳酸菌对活菌数的贡献较大有关。
2.4 STS测定结果
发酵糙米乳的STS与其黏性有关,黏性较好的发酵糙米乳对其组织状态有较好的保持作用。在未添加稳定剂或增稠剂的前提下,乳清析出较少或不析出,则感官品质较好,易于为消费者接受。随着pH值的下降,发酵糙米乳的STS下降,即发酵糙米乳的持水性增强。这与Olson等[25]的研究结果相符合。不同组合的STS值存在差异,其中Lp-Lr-La组合在发酵结束时高于其他组合大于30%,但是后期STS值较小,21 d时降到13.67%;Lp-La-L c组合的STS值整体较低,21 d时下降到14.96%。
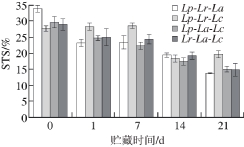
图3 复合乳酸菌发酵糙米乳STS变化
Fig. 3 Changes in STS during fermentation of brown rice milk by mixed LAB
2.5 蛋白质分解力
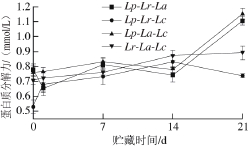
图4 复合乳酸菌发酵糙米乳蛋白质分解力变化
Fig. 4 Change in proteolytic activity during fermentation of brown rice milk by mixed LAB
适度的蛋白质分解力对于改善发酵糙米乳的质地和风味不可或缺,并且多菌种之间的互利共生作用也需要适当的蛋白质分解[26]。吴琼[27]以复合菌种发酵豆乳发现小分子量的蛋白条带增加,复合菌产生的蛋白酶作用于大分子蛋白质使其降解;小肽、氨基酸含量也明显增加,有利于人体的消化和吸收。冷藏期间氨基酸的含量呈增加的趋势,但是4 种组合的变化趋势并不一致:Lp-Lr-La组合曲折上升,Lp-Lr-Lc和Lp-La-Lc组合先上升后下降,Lr-La-Lc组合一直在缓慢上升。其中Lp-Lr-La、Lp-La-Lc和Lr-La-Lc组合的氨基酸含量较高,平均值约0.81 mmol/L,Lp-La-Lc组合则高达0.86 mmol/L。氨基酸总量下降可能是因为菌种自身消耗的氨基酸要高于降解产生的氨基酸数量,贮藏后期菌体数量下降所致。结合前期研究的结果发现,多菌种发酵糙米乳要高于单菌发酵糙米乳的蛋白分解力[15],这与白凤翎[28]的研究结果一致,混合菌培养时的远胜于单一菌株培养时的蛋白质水解力。
2.6 黏弹性测定结果
2.6.1 剪切扫描结果

图5 复合乳酸菌发酵糙米乳剪切速率-表观黏度曲线
Fig. 5 Plot of apparent viscosity against shear rate for fermented brown rice milk
由图5可知,随着剪切速率的增大,发酵糙米乳表观黏度出现明显下降。在0.01~1 s-1之间表观黏度急速下降,1~150 s-1之间表观黏度下降趋于平缓。之后开始降速剪切,表观黏度随着剪切速率的降低而增大,但是并未恢复到最初的黏度,上行线和下行线都没有重合,形成了触变环。这4 种菌种组合的发酵糙米乳均为假塑性流体,Lr-La-Lc组合表现出了最强的剪切稀化特性,Lp-Lr-
La和Lp-La-Lc组合的剪切稀化作用较弱,触变环面积分别为500.98、539.25 Pa/s。说明这2 组发酵糙米乳经外力作用后,黏度变化小,去除外力后发酵糙米乳恢复到未经外力作用时的状态所需时间较短,反映出样品经过长时间剪切再静止这一重新稠化的过程。此外与单菌种发酵糙米乳相比,触变环面积减小[15],也说明复合乳酸菌发酵糙米乳的结构破坏后更容易恢复。上述结果表明,复合菌种发酵糙米乳的流变学性质要优于单一菌种发酵糙米乳。
2.6.2 频率扫描结果
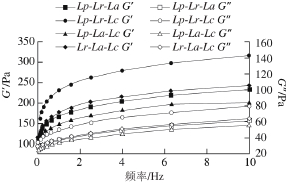
图6 复合乳酸菌发酵糙米乳G和G随频率的变化
Fig. 6 Plots of storage modulus and loss modulus against frequency for fermented brown rice milk
贮藏模量(G)衡量的是发酵糙米乳的弹性特征,损失模量(G)衡量的是发酵糙米乳的黏性特征。如图6所示,在0.1~10 Hz的频率扫描范围内,随着扫描频率的增大,G和G都增大。4 种发酵糙米乳的G都高于G,样品中弹性特征占优势。Lp-Lr-Lc组合的G和G均为最高,另外3 组G和G值相近。不同的G值反映了酸奶凝胶特性的不同[29],G值较小即弹性较小,凝胶结构相对疏松。与单菌株发酵糙米乳相比,其弹性、黏性特征均减小[15],可能是乳酸菌之间的相互作用使其黏弹性特征发生变化。
2.6.3 流变参数拟合结果
表2 复合乳酸菌发酵糙米乳稠度系数的变化情况
Table 2 Changes in consistency coeffi cient during fermentation of brown rice milk
Pa·sn
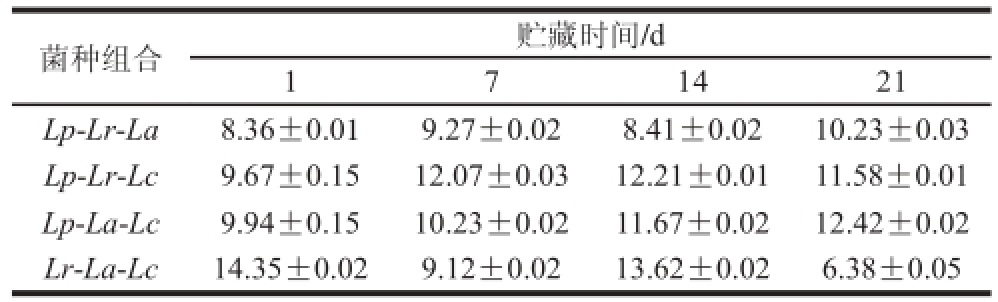
表3 复合乳酸菌发酵糙米乳流动行为指数的变化情况
Table 3 Change in fi ow behavior index during fermentation of brown rice milk
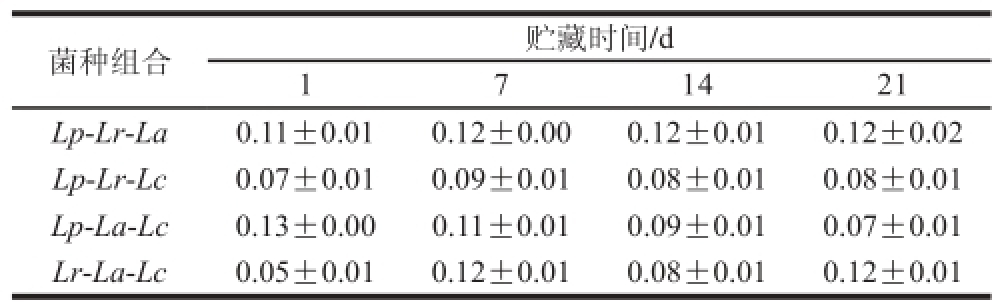
4 种样品的R值均高于0.99,说明该模型能够较好地拟合发酵糙米乳降速剪切时的流变学性质。如表2、3所示,发酵糙米乳的稠度系数和流动行为指数并未出现显著性差异,但是Lp-Lr-Lc和Lp-La-Lc组合的稠度系数较高,说明贮藏期间这2 组样品的稠度更大,Lp-Lr-La和Lp-La-Lc组合的流动行为指数较高,说明贮藏期间这2 组样品的剪切稀化作用不明显。
各菌种组合发酵的发芽糙米乳中,各菌种组合随着贮藏时间的增加表现出的趋势性变化较为显著,其中Lp-La-Lc组合在4 ℃贮藏21 d后,pH值下降了0.71 个单位,滴定酸度增加了29.62 ºT,是后酸化程度最弱的组合。随着贮藏时间的延长活菌数呈下降趋势,但数量一直较高,其中Lp-Lr-Lc和Lp-La-Lc组合是活菌数较高的两组,贮藏期内高达9(lg(CFU/g)),至21 d时降到8.7(lg(CFU/g))。4 组发酵糙米乳的STS值均呈下降的趋势,其中Lp-La-Lc组合的STS值整体较低,21 d时下降到14.96%,Lp-Lr-La组合在21 d时下降到13.67%。4 组发酵糙米乳的氨基酸含量均值约0.81 mmol/L,其中Lp-La-Lc组合较高为0.86 mmol/L。Lp-Lr-La和Lp-La-Lc组合的剪切稀化作用较弱,触变环面积分别为500.98、518.03 Pa/s,对发酵糙米乳的下行线进行拟合后得出Lp-Lr-La和Lp-La-Lc组合在21 d的贮藏期内流动行为指数较高,即剪切稀化作用弱。综合考虑,可以得出Lp-La-Lc这一乳酸菌菌种组合较为适合发酵发芽糙米乳。
参考文献:
[1] 张泓, 刘玉芳, 许杨. 发芽糙米的开发价值[J]. 食品工业科技, 2009, 30(6): 368-370. DOI:10.13386/j.issn1002-0306.2009.06.091.
[2] 王立, 段维, 钱海峰, 等. 糙米食品研究现状及发展趋势[J]. 食品与发酵工业, 2016, 42(2): 236-243. DOI:10.13995/j.cnki.11-1802/ ts.201602041.
[3] CHO D H, LIM S T. Germinated brown rice and its bio-functional compounds[J]. Food Chemistry, 2016, 196: 259-271. DOI:10.1016/ j.foodchem.2015.09.025.
[4] 房克敏, 李再贵, 袁汉成, 等. HPLC法测定发芽糙米中γ-氨基丁酸含量[J]. 食品科学, 2006, 27(4): 208-211.
[5] 程威威, 周婷, 吴跃, 等. 高效液相色谱法测定发芽糙米中γ-氨基丁酸含量[J]. 食品科学, 2014, 35(12): 98-101. DOI:10.7506/spkx1002-6630-201412019.
[6] 胡玲玲, 李春阳, 曾晓雄, 等. 富硒发芽糙米蛋白的抗氧化活性[J]. 食品科学, 2016, 37(1): 99-103. DOI:10.7506/spkx1002-6630-201601018.
[7] CHEN H H, CHANG H C, CHEN Y K, et al. An improved process for high nutrition of germinated brown rice production: low-pressure plasma[J]. Food Chemistry, 2016, 191: 120-127. DOI:10.1016/ j.foodchem.2015.01.083.
[8] ITO Y, SHEN M, KISE M, et al. Effect of pre-germinated brown rice on postprandial blood glucose and insulin level in subjects with hyperglycemia[J]. Japanese Journal of Food Chemistry, 2005, 12(2): 80-84.
[9] IMAM M U, ISHAKA A, OOI D J, et al. Germinated brown rice regulates hepatic cholesterol metabolism and cardiovascular disease risk in hypercholesterolaemic rats[J]. Journal of Functional Foods, 2014, 8: 193-203. DOI:10.1016/j.jff.2014.03.013.
[10] 李飞, 隋新, 毕丽娜, 等. 发芽糙米降血糖作用的研究[J]. 中国食品添加剂, 2016, 26(5): 168-171. DOI:10.3969/j.issn.1006-2513.2016.05.022.
[11] 孙月娥, 王卫东, 李曼曼. 发芽糙米黑豆复合饮料的生产工艺[J]. 食品科学, 2010, 31(18): 476-478.
[12] 李次力. 速食发芽糙米的研究[J]. 食品工业科技, 2010, 23(1): 304-306. DOI:10.13386/j.issn1002-0306.2010.01.023.
[13] COMEJO F, CACERES P J, ROSELL C M, et al. Effects of germination on the nutritive value and bioactive compounds of brown rice breads[J]. Food Chemistry, 2015, 173: 298-304. DOI:10.1016/ j.foodchem.2014.10.037.
[14] 唐明礼, 王勃, 刘贺, 等. 发芽糙米煎饼的研制[J]. 食品工业科技, 2015, 36(2): 261-270. DOI:10.13386/j.issn1002-0306.2015.02.048.
[15] 陈妍婕, 朱力杰, 何余堂, 等. 四种乳酸杆菌在全谷物糙米乳中发酵特性的研究[J]. 食品工业科技, 2016, 37(11): 179-184. DOI:10.13386/ j.issn1002-0306.2016.11.029.
[16] 陈海旭, 赵丽芹, 贠婷婷, 等. 益生菌发酵糙米饮料的制作工艺及其营养价值研究[J]. 食品与发酵工业, 2014, 40(11): 269-275. DOI:10.13995/j.cnki.11-1802/ts.201411047.
[17] 国家卫生和计划生育委员会. 食品酸度的测定: GB 5009.239—2016[S]. 北京: 中国标准出版社, 2016: 2-6.
[18] 曾凤泽. 具有优良发酵特性的德氏乳杆菌保加利亚亚种的筛选[D].呼和浩特: 内蒙古农业大学, 2013: 12.
[19] ABU-TATRBOUSH H M, AL-DAGAL M M, AL-ROYLI M A. Growth viability and proteolytic activity of bifidobacteria in whole camel milk[J]. Journal of Dairy Science, 1998, 81(2): 354-361. DOI:10.3168/jds.S0022-0302(98)75584-5.
[20] NIELSEN P M, PETERSEN D, DAMBMANN C. Improved method for determining food protein degree of hydrolysis[J]. Food Chemistry and Toxicology, 2001, 66(5): 642-646. DOI:10.1111/j.1365-2621.2001.tb04614.x.
[21] 孙懿琳. 弱后酸化保加利亚乳杆菌菌株的筛选及其后酸化机理[D].哈尔滨: 东北农业大学, 2013: 33.
[22] BEAL C, LOUVET P, CORRIEU G. Influence of controlled pH and temperature on the growth and acidification of pure cultures of Streptococcus thermophilus 404 and Lactobacillus bulgaricus 398[J]. Applied Microbiology and Biotechnology, 1989, 32(2): 148-154. DOI:10.1007/BF00165879.
[23] 韩俊华. 嗜酸乳杆菌部分益生特性的研究[C]//中国乳业科技大会论文集. 北京: 中国乳业科技组委会, 2002: 93-95.
[24] VINDEROLA C G, BAILO N, REINHEIMER J A. Survival of probiotic microflora in Argentinean yoghurts during refrigerated storage[J]. Food Research International, 2000, 33(2): 97-102. DOI:10.1016/S0963-9969(00)00011-9.
[25] OLSON D W, ARYANA K J. An excessively high Lactobacillus acidophilus inoculation level in yogurt lowers product quality during storage[J]. LWT-Food Science and Technology, 2008, 41(5): 911-918. DOI:10.1016/j.lwt.2007.05.017.
[26] NEVIANI E, GIRAFFA G, BRIZZI A, et al. Amino acid requirements and peptidase activities of Streptococcus salivarius subsp. thermophilus[J]. Journal of Applied Bacteriology, 1995, 79(3): 302-307. DOI:10.1111/j.1365-2672.1995.tb03141.x.
[27] 吴琼. 复合菌种发酵豆乳营养成分变化的研究[D]. 长春: 吉林农业大学, 2013: 32.
[28] 白凤翎. 蛋白水解物促乳酸菌增殖及高密度培养体系研究[D]. 北京: 北京林业大学, 2010: 118.
[29] DONKOR O N, HENRIKSSON A, VASILJEVIC T, et al. Rheological properties and sensory characteristics of set-type soy yogurt[J]. Journal of Agricultural and Food Chemistry, 2007, 55(24): 9868-9876. DOI:10.1021/jf071050r.
Fermentation Characteristics of Mixed Cultures of Lactic Acid Bacteria for the Production of a Germinated Brown Rice-Based Fermented Beverage
ZHU Lijie, CHEN Ning, CHEN Yanjie, LIU Xiuying, WANG Bo, LIU He, HE Yutang, MA Tao*
(National & Local Joint Engineering Research Center of Storage, Processing and Safety Control Technology for Fresh Agricultural and Aquatic Products, College of Food Science and Technology, Bohai University, Jinzhou 121013, China)
Abstract:Ternary mixed starter cultures of Lactobacillus plantarum, L. rhamnosus, L. acidophilus and L. casei were used for the fermentation of germinated brown rice milk. The best starter culture was determined by evaluating acidity, viable cell count, syneresis susceptibility, proteolytic activity and rheological property after culture at 4 ℃ for 21 days. The results showed that germinated brown rice milk fermented by a starter culture consisting of L. plantarum, L. acidophilus and L. casei displayed good quality, and exhibited weak post-acidification during the subsequent 21 days of storage at 4 ℃, with the pH being reduced by only 0.71. Additionally, the viable cell count changed somewhat during the refrigerated storage, remaining higher than 8.7 (lg(CFU/mL)), and the average content of free amino acids was up to 0.86 mmol/L. The rheological measurement indicated that its shear thinning behavior was weaker. In consequence, the co-culture of L. plantarum, L. acidophilus and L. casei was suitable for the fermentation of germinated brown rice milk.
Key words:Lactobacillus; co-culture; germinated brown rice; properties
DOI:10.7506/spkx1002-6630-201708014
中图分类号:TS213.4
文献标志码:A
文章编号:1002-6630(2017)08-0080-06
引文格式:
朱力杰, 陈宁, 陈妍婕, 等. 复合乳酸菌在发芽糙米乳中的发酵特性[J]. 食品科学, 2017, 38(8): 80-85. DOI:10.7506/ spkx1002-6630-201708014. http://www.spkx.net.cn
ZHU Lijie, CHEN Ning, CHEN Yanjie, et al. Fermentation ch aracteristics of mixed cultures of lactic acid bacteria for the production of a germinated brown rice-based fermented beverage[J]. Food Science, 2017, 38(8): 80-85. (in Chinese with English abstract)
DOI:10.7506/spkx1002-6630-201708014. http://www.spkx.net.cn
收稿日期:2016-06-15
基金项目:辽宁省农业领域青年科技创新人才培养计划项目(2015001)
作者简介:朱力杰(1986—),男,讲师,博士,研究方向为农产品深加工与功能性食品。E-mail:lijiezhu325@126.com
*通信作者:马涛(1962—),男,教授,博士,研究方向为粮油与植物蛋白工程。E-mail:matao-09@163.com