
高 星1,2,李 欣1,李 铮1,丁 武2,张德权1,*
(1.中国农业科学院农产品加工研究所,农业部农产品加工综合性重点实验室,北京 100193;
2.西北农林科技大学食品科学与工程学院,陕西 杨凌 712100 )
摘 要:目的:研究改变肌动球蛋白粗提物磷酸化水平对其解离程度及ATPase活性的 调控作用。方法:以肌动球蛋白粗提物为材料,添加蛋白激酶A(protein kinase A,PKA)和碱性磷酸酶(alkaline phosphatase,AP)改变蛋白的磷酸化水平(4 ℃孵育48 h),通过十二烷基硫酸钠-聚丙烯酰胺凝胶电泳和荧光染色、蛋白质免疫印迹、ATPase活力测定试剂盒测定蛋白的磷酸化水平、游离肌动球蛋白含量和肌动球蛋白ATPase活力随孵育时间的变化。结果:PKA处理组蛋白样品中的肌钙蛋白T和肌球蛋白调节轻链的磷酸化水平显著高于对照组(P<0.05),游离肌动蛋白含量显著高于对照组,且在0~2 h显著升高,2~48 h缓慢升高;肌动球蛋白ATPase活力显著低于对照组(P<0.05),其活力随孵育时间延长显著升高(P<0.05);AP处理组的变化则与之相反。结论:肌钙蛋白T、肌球蛋白调节轻链的磷酸化降低了肌球蛋白与肌动蛋白之间的相互作用力(肌动球蛋白ATPase活力低),从而促进肌动球蛋白的解离。
关键词:磷酸化;肌球蛋白调节轻链;肌钙蛋白T;肌动球蛋白解离
肌球蛋白和肌动蛋白是肌原纤维中含量最丰富的两种蛋白质,分别约占肌原纤维蛋白总量的50%和20%[1]。动物宰后由于ATP的丧失,促使肌动蛋白和肌球蛋白之间交联结合形成肌动球蛋白,而肌动球蛋白复合体的解离是影响肉品嫩度的重要因素之一,肌动球蛋白的解离程度越大,相互作用力越小,肉嫩度越好。磷酸化的肌球蛋白是维持细胞骨架活性及细胞功能的重要效应因子[2-3]。肌球蛋白调节轻链周期性地磷酸化与去磷酸化,是细胞发生运动和收缩的必备条件[4],对肌球蛋白与肌动蛋白的相互作用具有重要的调控作用[5-6]。1977年,Pires等[7]发现快肌肌球蛋白调节轻链的磷酸化作用可以改变肌球蛋白与肌动蛋白的相互作用。随后的研究表明,肌球蛋白调节轻链的磷酸化修饰会破坏肌肉粗丝和细丝的相互作用,使肌球蛋白重链头部远离粗丝[8-10],同时调控肌球蛋白的活性[11]和肌节的收缩[12],从而影响肉最终的嫩度。
本团队前期研究表明,宰后成熟过程中肌球蛋白调节轻链的磷酸化水平与肌动球蛋白解离具有很高的相关性,但是肌球蛋白调节轻链的磷酸化作用如何调控肌动球蛋白的解离,以及肌动-肌球蛋白相互作用(包括相互作用力和相互作用量)的变化是单纯由肌球蛋白调节轻链的磷酸化调控,还是肉中各种酶共同作用调控的,这些机理均有待深入研究。本研究以提取的羊背最长肌肌动球蛋白粗品为原料,通过添加蛋白激酶A(protein kinase A,PKA)和碱性磷酸酶(alkaline phosphatase,AP),制备高、低磷酸化水平样品,研究蛋白质磷酸化水平的改变对肌动球蛋白解离、肌动球蛋白ATPase活性的影响,进一步揭示肌原纤维蛋白磷酸化修饰对肌动-肌球蛋白相互作用的调控机制。
1.1 材料与试剂
本实验选取饲养条件相同的3 只10 月龄的小尾寒羊与大尾寒羊的杂交公羊,在北京市昌平区商业化的屠宰场屠宰,平均胴体质量约23 kg。取宰后30 min的羊背最长肌,剔除可见的筋膜及结缔组织,分装于2 g冻存管后立即用液氮速冻,于-80 ℃保存。
三羟甲基氨基甲烷(tris(hydroxymethyl)methyl aminomethane,THAM)、十二烷基硫酸钠(sodium dodecyl sulfate,SDS)、丙烯酰胺、甲叉双丙烯酰胺、过硫酸铵(ammonium persulphate,APS)、四甲基乙二胺(tetramethylethylenediamine,TEMED)、ATPase活力测定试剂盒、PKA、AP、肌动蛋白抗体、羊抗鼠二抗 美国Sigma公司;磷酸酶抑制剂 瑞士Roche公司;二喹啉甲酸(bicinchoninic acid,BCA)蛋白检测试剂盒美国Thermo Fisher Scientific公司;Pro-Q Diamond、 SYPRO Ruby染色液 美国Invitrogen公司;氯化钠、碳酸钠、碳酸氢钠(均为国产分析纯) 北京化学试剂公司。
1.2 仪器与设备
Ultra Turrax Disperser S25分散器 德国IKA公司;Himac CR 22 GII高效冷冻离心机 日本Hitachi公司;恒温振荡金属浴 杭州奥盛仪器有限公司;SpectraMax 190全波长酶标仪 美国Molecular Devices公司;电泳设备(Mini-PROTEAN Tetra System)、半干转膜设备(Trans-Blot TurboTM)、ChemiDocTMMP凝胶成像系统美国Bio-Rad 公司。
1.3 方法
1.3.1 样品制备
肌动球蛋白粗提物的制备参考Okitani等[13]的方法,并稍作修改:取1 g肉,加入10 mL Weber-Edsall溶液(0.6 mol/L KCl、0.04 mol/L NaHCO3、0.01 mol/L Na2CO3,pH 7.2),进行匀浆。匀浆液置于4℃培养箱内,摇床振荡24 h后,用两层尼龙网过滤除去不溶物质,然后加入超纯水使提取液中K+浓度稀释至0.2 mol/L,4 ℃条件下摇床振荡60 min,离心(4 ℃、15 000×g、20 min)。取沉淀溶解于3 mL KCl-Tris溶液中(0.6 mol/L KCl、20 mmol/L Tris-HCl、pH 7.2),即为肌动球蛋白粗提物溶液。
用BCA试剂盒测定蛋白质量浓度,将蛋白质量浓度调至4 mg/mL,同时使蛋白溶液含有5 mmol/L ATP、10 mmol/L MgCl2、1 mmol/L CaCl2,以保证后续酶反应的正常进行。将所得的匀浆液平均分为3 份,分别添加AP(1 U/4 μg pro)、PKA(1 U/10 μg pro)以及等量的超纯水,在4 ℃条件下振荡孵育2、12、24、48 h。在每个时间点每份样品平均分为3 份,一份添加2 倍体积的上样缓冲液在沸水中煮沸5 min,得到样品Ⅰ,用于蛋白磷酸化水平的测定。一份添加两倍体积的超纯水使溶液中KCl的浓度变为0.2 mol/L,然后离心(4 ℃、15 000×g、20 min),取上清液(包含游离肌动蛋白)添加等量的上样缓冲液煮沸5 min,所得的样品Ⅱ用于测定肌动球蛋白解离程度。另一份即为样品Ⅲ,液氮速冻后于-80 ℃保存,用于测定肌动球蛋白ATPase活力。
1.3.2 蛋白磷酸化水平的分析
将样品Ⅰ离心(12 000×g、1 min),取上清液上样,上样量为7 μL(蛋白质量浓度为1.3 μg/μL)。在70 V电压下开始电泳,结束后进行Pro-Q染色和Ruby染色,染色流程参考陈立娟等[14]的方法。
使用ChemiDocTMMP凝胶成像系统对染色后的凝胶拍照,利用Quantity One 4.6.2软件对肌钙蛋白T和肌球蛋白调节轻链两条条带的光密度值进行定量,得到对应的Pro-Q Diamond染色(磷酸化蛋白染色)光密度值(P)和SYPRO Ruby染色(全蛋白染色)光密度值(T),该条带的光密度值之比P/T即为该条带蛋白的蛋白质磷酸化水平。
1.3.3 肌动球蛋白解离程度的测定
将样品Ⅱ离心(12 000×g、1 min),取上清液上样,上样量均为10 μL(蛋白质量浓度为0.65 μg/μL)。电泳结束后用半干转膜仪将凝胶中的蛋白转印到聚偏氟乙烯(polyvinylidene fluoride,PVDF)膜上,TBS溶液(0.01 mol/L Tris、0.15 mol/L NaCl,pH 7.5)漂洗(3 次,2 min/次)后,用封闭液(含0.05% Tween-20、3%牛血清白蛋白的TBS溶液)室温条件下封闭2 h。然后与肌动蛋白抗体(用封闭液1∶1 000稀释)室温孵育2 h,随后将膜用TBST1溶液(含0.1% Tween-20的TBS溶液)漂洗3 次后与羊抗鼠二抗(用封闭液1∶5 000稀释)室温孵育2 h。膜用TBST2溶液(0.05 mol/L Tris、0.15 mol/L NaCl、0.1% Tween-20,pH 7.5)漂洗3 次后使用电化学发光(electro chemiluminescence,ECL)显色剂显色,并使用凝胶成像仪拍照。采用Quantity One软件分析蛋白免疫印迹条带光密度值,用得到的游离肌动蛋白光密度值的大小表示肌动球蛋白的解离程度,游离肌动蛋白光密度值大表明肌动球蛋白解离程度高,反之则低。
1.3.4 肌动球蛋白ATPase活力的测定
肌动球蛋白ATPase活力的测定按照ATPase活力测定试剂盒的说明书进行测定。由式(1)计算出样品吸光度的变化,由式(2)计算出样品中无机磷的含量([Pi]),由式(3)计算出肌动球蛋白ATPase活力。
式中:Vr为反应体积(40 μL);Vs为样品体积(10 μL);t为反应时间/min;酶活力单位U定义为实验条件下每分钟催化生成1 μmol无机磷所需要的酶的量。
1.4 统计分析
所有实验均做3 次重复,每个样品做3 次平行,所得数据采用SPSS 19.0统计软件进行方差分析,通过最小显著差异法(least signif i cant difference,LSD)和邓肯氏总重比较法(Duncans Multiple-rang test)进行差异显著性分析。
2.1 不同酶处理对蛋白质磷酸化水平的影响
对PKA、AP和对照3 个处理组孵育不同时间(0、2、12、24、48 h)的样品进行SDS-聚丙烯酰胺凝胶电泳(polyac rylamide gel electrophoresis,PAGE)后采用磷酸化染色(图1A)与全蛋白染色(图1B)进行分析,结果表明不同处理组之间蛋白样品中只有部分条带的灰度值发生了明显的变化。磷酸化染色凝胶图中条带1(肌钙蛋白T)和条带2(肌球蛋白调节轻链)的PKA处理组的灰度值强度明显强于对照组,而AP处理组的灰度值强度则明显弱于对照组,这说明PKA和AP可以有效地作用于样品中的部分蛋白,导致其磷酸化水平发生改变。本实验所处理的样品为肌动球蛋白粗提物,其中不含有酶类,因此对照组中样品蛋白的磷酸化水平随孵育时间基本没有发生变化。

图1 不同处理组孵育不同时间样品磷酸化蛋白与全蛋白染色效果
Fig.1 SDS-PAGE images of phosphoproteins and total proteins for different groups at different incubation times

图2 不同处理组孵育不同时间肌钙蛋白T的磷酸化水平
Fig.2 Variation in troponin T phosphorylation level in different treatments with incubation time
对图1电泳图中的条带1和条带2进行灰度值分析,用磷酸化蛋白染色(图1A)的光密度值与全蛋白染色(图1B)的光密度值之比表示磷酸化水平,分别得到肌钙蛋白T(图2)和肌球蛋白调节轻链(图3)的磷酸化水平的变化趋势图。由图2可知,对照组中肌钙蛋白T的磷酸化水平随孵育时间基本没有变化,AP处理组中肌钙蛋白T的磷酸化水平在0~2 h之间显著下降(P<0.05),随后在2~48 h之间维持稳定,说明在本实验条件下AP在2 h内就可以有效地作用于肌钙蛋白T,使其发生去磷酸化反应。而PKA处理组中肌钙蛋白T的磷酸化水平在0~2 h间不断上升(P<0.05),随后则维持稳定(12~48 h),说明PKA可以有效地作用于样品蛋白中的肌钙蛋白T,促使其发生磷酸化反应,使磷酸化水平升高。在同一时间点AP组肌钙蛋白T磷酸化水平显著低于对照组(P<0.05),PKA组肌钙蛋白T磷酸化水平显著高于对照组(P<0.05)。

图3 不同处理组孵育不同时间肌球蛋白调节轻链的磷酸化水平
Fig.3 Variation in myosin regulatory light chain phosphorylation level in different treatments with incubation time
如图3所示,AP和PKA促使肌球蛋白调节轻链发生去磷酸化或磷酸化反应,降低或提高肌球蛋白调节轻链的磷酸化水平。在AP处理组肌球蛋白调节轻链的磷酸化水平在0~12 h之间没有显著变化,在12~48 h之间显著下降(P<0.05),而PKA处理组中肌球蛋白调节轻链的磷酸化水平在0~12 h之间没有显著变化,在12~48 h间显著升高(P<0.05)。不同处理组之间肌球蛋白调节轻链磷酸化水平在12 h没有显著差异,而在其他时间点AP处理组肌球蛋白调节轻链磷酸化水平显著低于对照组(P<0.05),PKA处理组肌球蛋白调节轻链磷酸化水平显著高于对照组(P<0.05)。
2.2 不同酶处理对肌动球蛋白解离程度的影响
利用免疫印迹测定AP、对照和PKA 3 个处理组孵育不同时间的游离肌动蛋白的量的变化,其结果如图4所示。图中对照和PKA处理组中的游离肌动蛋白量在0~2 h显著升高,2~48 h缓慢升高,AP处理组游离肌动蛋白量随孵育时间显著下降。在同一时间点AP处理组游离肌动蛋白条带的灰度值低于对照组,PKA处理组则高于对照组。这说明在4 ℃孵育条件下,肌动球蛋白发生解离,而促进蛋白的去磷酸化作用可以抑制肌动球蛋白的解离,促进蛋白的磷酸化作用促进肌动球蛋白的解离。采用ChemiDocTMMP凝胶成像系统对图像进行ECL显色曝光,并采用Image Lab软件获取曝光图像。在软件自动曝光条件下,PKA和对照处理组的游离肌动蛋白条带信号灵敏度高,而AP处理组则由于样品中所含的游离肌动蛋白量太少,低于检测限,图中没有条带(图4A)。将曝光时间延长至5 s时,在AP处理组可以看到较浅的肌动蛋白条带,该处理组肌动蛋白条带的灰度值明显低于对照组和PKA处理组(图4B)。
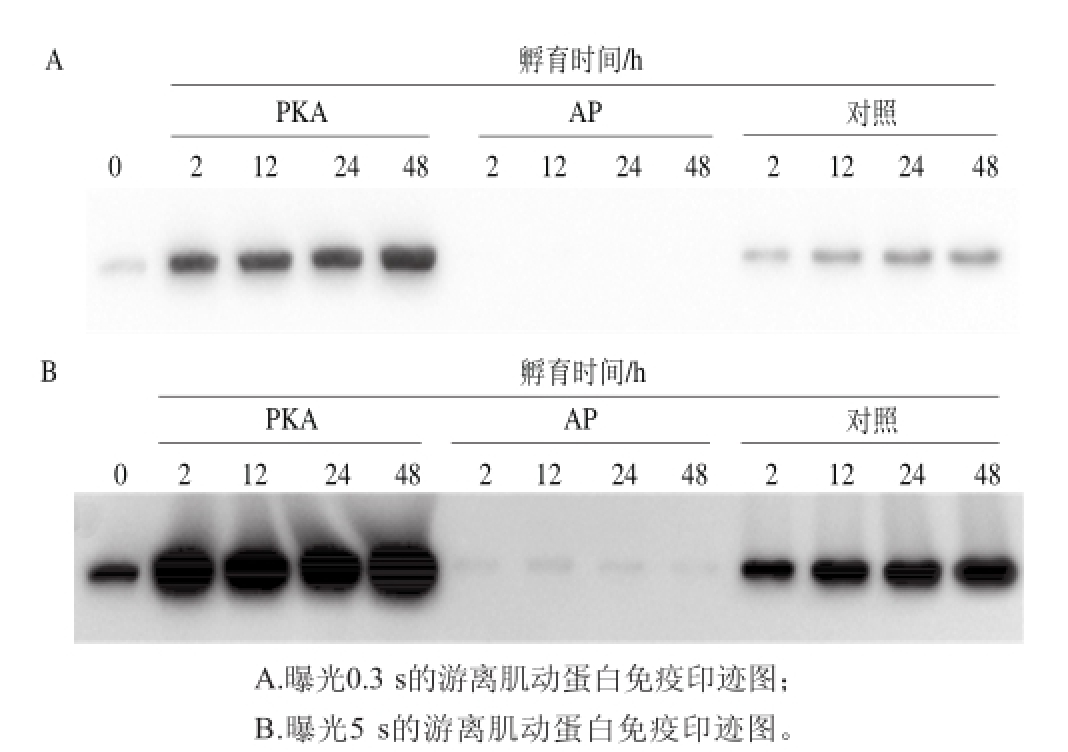
图4 不同处理组孵育不同时间游离肌动蛋白量的变化
Fig.4 Variation in actin contents in different treatments with incubation time
2.3 不同酶处理对肌动球蛋白ATPase活性的影响
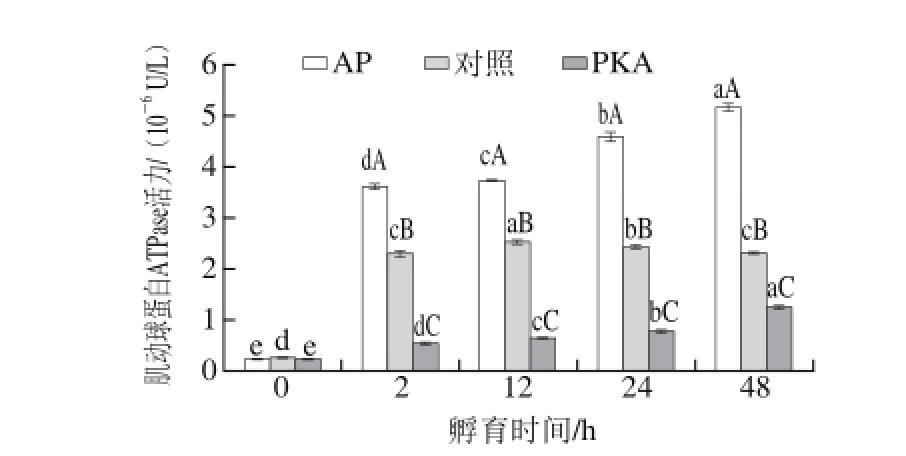
图5 不同处理组孵育不同时间肌动球蛋白ATPase活性的变化
Fig.5 Variation in actomyosin ATPase activity in different treatments with incubation time
肌动球蛋白ATPase活性通常用来表示肌球蛋白和肌动蛋白的相互作用,它的大小表示肌动球蛋白复合体的完整性[15-16],肌动球蛋白ATPase活性减小表示肌球蛋白-肌动蛋白相互作用力的减弱。AP、对照和PKA 3 个处理组孵育不同时间的肌动球蛋白ATPase活力的变化如图5所示。AP和PKA处理组的肌动球蛋白ATPase活力随孵育时间呈现出逐渐增大的趋势(P<0.05),而对照组的肌动球蛋白ATPase活力在0~12 h之间逐渐增大(P<0.05),12~48 h之间则逐渐减小(P<0.05)。在同一时间点,不同的处理组之间肌动球蛋白ATPase活力差异显著,AP处理组的肌动球蛋白ATPase活力均显著高于对照组(P<0.05),而PKA处理组的肌动球蛋白ATPase活力显著低于对照组(P<0.05)。
骨骼肌肌动球蛋白的解离受多种因素的影响,如Ca2+浓度、ATP及其水解产物、内源酶、多种相关蛋白及蛋白磷酸化作用等。在体内,当Ca2+与肌钙蛋白C结合后解除了肌钙蛋白I、肌钙蛋白T和原肌球蛋白对肌动蛋白-肌球蛋白相互作用的抑制作用,负荷ATP的肌球蛋白横桥就立即与肌动蛋白结合形成肌动球蛋白,这时横桥中的ATPase被激活,分解ATP,释放出ADP和无机磷酸,并释放出能量,使横桥发生扭转,促使肌节收缩[17],接着肌动蛋白接受另一个ATP分子,促使横桥断裂[1]。本研究结果表明,肌钙蛋白T磷酸化水平升高,肌动球蛋白ATPase活性显著降低,肌动球蛋白解离程度显著增大;肌钙蛋白T磷酸化水平降低,肌动球蛋白ATPase活性显著升高,肌动球蛋白解离程度显著降低。有研究表明心肌肌钙蛋白T的磷酸化作用可以影响肌肉收缩[18]。肌钙蛋白T的磷酸化作用降低了它与原肌球蛋白的亲和力,从而使肌球蛋白结合位点暴露出来,降低Ca2+依赖性的肌动球蛋白ATPase活性[19-20],同时改变了其降解的灵敏性[21-22],导致肌动球蛋白易于发生解离[23],这与本研究中的结果相一致。Noland[24]、Wattanapermpool[25]等用PKA处理无皮纤维,结果发现PKA可以降低细丝的Ca2+敏感度,从而改变横桥作用力和ATPase活性。其他研究也发现PKA处理可以降低细丝的Ca2+亲和力,而且这是导致横桥数量减少的直接影响因素[26]。因此,PKA调控肌原纤维蛋白的磷酸化作用,加速肌丝舒张[27-28],直接影响肌球蛋白横桥的动力转换[29]。
肌球蛋白调节轻链在体内可以发生可逆的磷酸化及去磷酸化反应[30-31]。肌球蛋白调节轻链磷酸化后肌球蛋白头部的构象发生改变,导致肌球蛋白两个头部的相互作用发生变化,从而调节肌动球蛋白的解离与ATPase活性[32-33]。在平滑肌中,肌球蛋白调节轻链的磷酸化是一种主要的调节机制,它促使肌球蛋白横桥从粗丝表面释放出来。然而,在哺乳动物横纹肌中,肌球蛋白调节轻链的磷酸化只是一种辅助调节机制[34],未磷酸化的肌球蛋白调节轻链所在的肌球蛋白头部与肌粗丝松散的连接在一起,肌球蛋白调节轻链的磷酸化作用降低了这种连接作用。因此,肌球蛋白调节轻链磷酸化的主要作用是促使肌球蛋白横桥头部远离肌粗丝,而不仅仅是改变肌球蛋白头部的运动性[35]。本研究中,当肌球蛋白调节轻链磷酸化水平升高时,肌动球蛋白ATPase活性显著降低,肌动球蛋白解离程度显著增大;而当肌球蛋白调节轻链磷酸化水平降低时,肌动球蛋白ATPase活性显著升高,肌动球蛋白解离程度显著降低。这说明肌球蛋白调节轻链的磷酸化与去磷酸化反应调节肌球蛋白与肌动蛋白之间相互作用力的减弱或增强,从而促使肌动球蛋白解离程度的增大或降低。同时,Oda[5]和Wagner[6]等研究发现肌球蛋白调节轻链结构的改变还可以影响肌球蛋白头部与肌动蛋白的相互作用,磷酸化与去磷酸化的肌球蛋白的协同作用机制促使去磷酸化的肌球蛋白再次与肌动蛋白结合[36-37]。
另外,本研究中ATP不仅是磷酸化反应发生的必要条件,也是影响肌动-肌球蛋白相互作用的一个重要因素。Okitani等[13,38]将鸡、猪、牛骨骼肌中的肌动球蛋白在0.2 mol/L KCl的溶液中(0 ℃、pH 7.2)孵育,结果发现ATP的水解产物如腺苷酸(adenosine monophosphate,AMP)或肌苷酸(inosincacid,IMP)等会促使肌动球蛋白发生不可逆的解离,分解为肌动蛋白和肌球蛋白。
PKA和AP可以有效地促使肌钙蛋白T和肌球蛋白调节轻链发生磷酸化或去磷酸化反应,提高或降低肌球蛋白调节轻链的磷酸化水平。随着孵育的进行,PKA处理组中肌钙蛋白T和肌球蛋白调节轻链的磷酸化水平逐渐升高,肌动球蛋白的解离程度增大,肌动球蛋白ATPase活性降低。而AP处理组中肌钙蛋白T和肌球蛋白调节轻链的磷酸化水平降低,肌动球蛋白的解离程度减小,肌动球蛋白ATPase活性显著升高。由此可知,肌钙蛋白T与肌球蛋白调节轻链的磷酸化/去磷酸化与肌动球蛋白解离程度和ATPase活性紧密相关。高磷酸化水平组肌动-肌球蛋白的相互作用力显著减弱(肌动球蛋白ATPase活性小),肌动球蛋白解离程度增大;而低磷酸化水平组肌动-肌球蛋白的相互作用力显著增大,肌动球蛋白解离程度减小。
参考文献:
[1] 尹靖东. 动物肌肉生物学与肉品科学[M]. 北京: 中国农业大学出版社, 2011: 64-67.
[2] CHARDIN P. GTPase regulation: getting aRnd Rock and Rho inhibition[J]. Current Biology, 2003, 13(18): R702-R704. DOI:10.1016/j.cub.2003.08.042.
[3] NAKAMURA A, XIE C, ZHANG Y, et al. Role of non-kinase activity of myosin light-chain kinase in regulating smooth muscle contraction, a review dedicated to Dr. Setsuro Ebashi[J]. Biochemical and Biophysical Research Communications, 2008, 369(1): 135-143. DOI:10.1016/j.bbrc.2007.11.096.
[4] MATSUMURA F, HARTSHORNE D J. Myosin phosphatase target subunit: many roles in cell function[J]. Biochemical and Biophysical Research Communications, 2008, 369(1): 149-156. DOI:10.1016/ j.bbrc.2007.12.090.
[5] ODA S, ORIOL-AUDIT C, REISLER E. Effects of actin and calcium ion on chymotryptic digestion of skeletal myosin and their implications to the function of light chains[J]. Biochemistry, 1980, 19(24): 5614-5618. DOI:10.1021/bi00565a024.
[6] WAGNER P D. Effect of skeletal muscle myosin light chain 2 on the Ca2+-sensitive interaction of myosin and heavy meromyosin with regulated actin[J]. Biochemistry, 1984, 23(25): 5950-5956. DOI:10.1021/bi00320a010.
[7] PIRES E M, PERRY S V. Purif i cation and properties of myosin lightchain kinase from fast skeletal muscle[J]. Biochemical Journal, 1977, 167(1): 137-146. DOI:10.1042/bj1670137.
[8] ALAMO L, WRIGGERS W, PINTO A, et al. Three-dimensional reconstruction of tarantula myosin filaments suggests how phosphorylation may regulate myosin activity[J]. Journal of Molecular Biology, 2008, 384(4): 780-797. DOI:10.1016/j.jmb.2008.10.013.
[9] BRITO R, ALAMO L, LUNDBERG U, et al. A molecular model of phosphorylation-based activation and potentiation of tarantula muscle thick fi laments[J]. Journal of Molecular Biology, 2011, 414(1): 44-61. DOI:10.1016/j.jmb.2011.09.017.
[10] SHEIKH F, OUYANG K F, CAMPBELL S G, et al. Mouse and computational models link Mlc2v dephosphorylation to altered myosin kinetics in early cardiac disease[J]. The Journal of Clinical Investigation, 2012, 122(4): 1209-1221. DOI:10.1172/JCI61134.
[11] KASZA K E, FARRELL D L, ZALLEN J A. Spatiotemporal control of epithelial remodeling by regulated myosin phosphorylation[J]. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(32): 11732-11737. DOI:10.1073/pnas.
[12] RASSIER D E. The effects of length on fatigue and twitch potentiation in human skeletal muscle[J]. Clinical Physiology, 2000, 20(6): 474-482.
[13] OKITANI A, ICHINOSE N, ITOH J, et al. Liberation of actin from actomyosin in meats heated to 65 degrees C[J]. Meat Science, 2009, 81(3): 446-450. DOI:10.1016/j.meatsci.2008.09.008.
[14] 陈立娟, 李欣, 杨扬, 等. 不同嫩度羊肉肌浆蛋白质磷酸化水平随宰后成熟时间变化的研究[J]. 现代食品科技, 2015, 31(4): 95-101.
[15] OUALI A, VALIN C. Effect of muscle lysosomal enzymes and calcium activated neutral proteinase on myofibrillar ATPase activity: relationship with ageing changes[J]. Meat Science, 1981, 5(3): 233-245. DOI:10.1016/0309-1740(81)90006-1.
[16] PAREDI M, CRUPKIN M, Biochemical and physicochemical properties of actomyosin from pre- and post-spawned flounder (Paralichtys patagonicus) stored on ice[J]. LWT-Food Science and Technology, 2007, 40(10): 1716-1722. DOI:10.1016/j.lwt.2006.11.011.
[17] EBASHI S. Separation of troponin into its three components[J]. Journal of Biochemistry, 1972, 72(3): 787-790.
[18] STRENG A S, de BOER D, van der VELDEN J, et al. Posttranslational modifications of cardiac troponin T: an overview[J]. Journal of Molecular & Cellular Cardiology, 2013, 63: 47-56. DOI:10.1016/ j.yjmcc.2013.07.004.
[19] NOLAND T A, KUO J F. Protein kinase C phosphorylation of cardiac troponin I or troponin T inhibits Ca2+-stimulated actomyosin MgATPase activity[J]. Journal of Biological Chemistry, 1991, 266(8): 4974-4978. DOI:10.1006/jmcc.1993.1007.
[20] NOLAND T A, KUO J F. Protein kinase C phosphorylation of cardiac troponin T decreases Ca2+-dependent actomyosin Mg ATPase activity and troponin T binding to tropomyosin-F-actin complex[J]. Biochemical Journal, 1992, 288(1): 123-129. DOI:10.1042/bj2880123.
[21] MAZÓN M, GANCEDO J, GANCEDO C. Phosphorylation and inactivation of yeast fructose-bisphosphatase in vivo by glucose and by proton ionophores. A possible role for cAMP[J]. European Journal of Biochemistry, 1982, 127(3): 605-608. DOI:10.1111/j.1432-1033.1982. tb06915.x.
[22] MÜLLER D, HOLZER H. Regulation of fructose-1,6-bisphosphatase in yeast by phosphorylation/dephosphorylation[J]. Biochemical & Biophysical Research Communications, 1981, 103(3): 926-933. DOI:10.1016/0006-291X(81)90899-8.
[23] HÜNLICH M, BEGIN K, GORGA J, et al. Protein kinase A mediated modulation of acto-myosin kinetics[J]. Journal of Molecular & Cellular Cardiology, 2005, 38(1): 119-125. DOI:10.1016/j.yjmcc.2004.10.005.
[24] NOLAND T A, GUO X D, RAYNOR R L, et al. Cardiac troponin I mutants. Phosphorylation by protein kinases C and A and regulation of Ca2+-stimulated MgATPase of reconstituted actomyosin S-1[J]. Journal of Biological Chemistry, 1995, 270(43): 25445-25454. DOI:10.1074/ jbc.270.43.25445.
[25] WATTANAPERMPOOL J, GUO X D, SOLARO R J. The unique amino-terminal peptide of cardiac troponin I regulates myofibrillar activity only when it is phosphorylated[J]. Journal of Molecular & Cellular Cardiology, 1995, 27(7): 1383-1391. DOI:10.1006/ jmcc.1995.0131.
[26] ROBERTSON S, JOHNSON J, HOLROYDE M, et al. The effect of troponin I phosphorylation on the Ca2+-binding properties of the Ca2+-regulatory site of bovine cardiac troponin[J]. Journal of Biological Chemistry, 1982, 257(1): 260-263.
[27] SAEKI Y, KOBAYASHI T, MINAMISAWA S, et al. Protein kinase A increases the tension cost and unloaded shortening velocity in skinned rat cardiac muscle[J]. Journal of Molecular & Cellular Cardiology, 1997, 29(6): 1655-1663. DOI:10.1006/jmcc.1997.0401.
[28] STRANG K, MOSS R. Beta-adrenergic receptor stimulation increases unloaded shortening velocity of skinned single ventricular myocytes from rats[J]. Circulation Research, 1994, 74(3): 542-549.
[29] HOH J F, ROSSMANITH G H, KWAN L J, et al. Adrenaline increases the rate of cycling of crossbridges in rat cardiac muscle as measured by pseudo-random binary noise-modulated perturbation analysis[J]. Circulation Research, 1988, 62(3): 452-461. DOI:10.1016/0022-2828(90)91480-U.
[30] PERRIE W T, PERRY S V. An electrophoretic study of the lowmolecular-weight components of myosin[J]. Biochemical Journal, 1970, 119(1): 31-38. DOI:10.1042/bj1190031.
[31] PERRIE W T, SMILLIE L B, PERRY S B. A phosphorylated lightchain component of myosin from skeletal muscle[J]. Biochemical Journal, 1973, 135(1): 151-164. DOI:10.1042/bj1350151.
[32] PEMRICK S M. The phosphorylated L2 light chain of skeletal myosin is a modifier of the actomyosin ATPase[J]. Journal of Biological Chemistry, 1980, 255(18): 8836-8841.
[33] VIBERT P, COHEN C. Domains, motions and regulation in the myosin head[J]. Journal of Muscle Research and Cell Motility, 1988, 9(4): 296-305. DOI:10.1007/BF01773873.
[34] MORGAN M, PERRY S V, OTTAWAY J. Myosin light-chain phosphatase[J]. Biochemical Journal, 1976, 157(3): 687-697.
[35] SWEENEY H L, BOWMAN B F, STULL J T. Myosin light chain phosphorylation in vertebrate striated muscle: regulation and function[J]. American Journal of Physiology, 1993, 264: C1085- C1095.
[36] HIMPENS B, MATTHIJS G, SOMLYO A V, et al. Cytoplasmic free calcium, myosin light chain phosphorylation, and force in phasic and tonic smooth muscle[J]. Journal of General Physiology, 1988, 92(6): 713-729.
[37] VYAS T B, MOOERS S U, NARAYAN S R, et al. Cooperative activation of myosin by light chain phosphorylation in permeabilized smooth muscle[J]. American Journal of Physiology, 1992, 263: C210-C219.
[38] OKITANI A, ICHINOSE N, KOZA M, et al. AMP and IMP dissociate actomyosin into actin and myosin[J]. Bioscience Biotechnology & Biochemistry, 2008, 72(8): 2005-2011. DOI:10.1271/bbb.80128.
Effect of Phosphorylation on Actomyosin Dissociation
GAO Xing1,2, LI Xin1, LI Zheng1, DING Wu2, ZHANG Dequan1,*
(1. Key Laboratory of Agro-Products Processing, Ministry of Agriculture, Institute of Food Science and Technology, Chinese Academy of Agricultural Sciences, Beijing 100193, China; 2. College of Food Science and Engineering, Northwest A&F University, Yangling 712100, China)
Abstract:Objective: The objective of this study was to investigate the regulation of actomyosin dissociation and ATPase activity in crude actomyosin extract by changing the phosphorylation level of actomyosin. Methods: The crude actomyosin extract was incubated with protein kinase A and alkaline phosphatase at 4 ℃ for 48 h. Subsequently, the changes in protein phosphorylation, actomyosin dissociation and ATPase activity were measured by SDS-PAGE, Pro-Q Diamond phosphoprotein gel staining kit, Western blotting and ATPase activity assay kit. Results: The phosphorylation levels of troponin T (TnT) and myosin regulatory light chain (MRLC) were signif i cantly higher in the protein kinase A group compared with the control group. The amount of liberated actin in the protein kinase A group was signif i cantly higher than that in control group, and it increased signif i cantly during the fi rst two hours and increased slightly during the following 46 hours in both groups. In addition, the actomyosin ATPase activity decreased signif i cantly in the protein kinase A group compared with the control group, and it increased signif i cantly with incubation time in the two groups. However, the opposite changes were observed for the alkaline phosphatase group. Conclusion: The phosphorylation of TnT and MRLC reduces the binding force between myosin and actin and thereby promotes the dissociation of actomyosin.
Key words:phosphorylation; myosin regulatory light chain; troponin T; actomyosin dissociation
DOI:10.7506/spkx1002-6630-201709004
中图分类号:TS201.2
文献标志码:A
文章编号:1002-6630(2017)09-0021-06
引文格式:
高星, 李欣, 李铮, 等. 肌动球蛋白磷酸化对其解离的影响[J]. 食品科学, 2017, 38(9): 21-26. DOI:10.7506/spkx1002-6630-201709004. http://www.spkx.net.cn
GAO Xing, LI Xin, LI Zheng, et al. Effect of phosphorylation on actomyosin dissociation[J]. Food Science, 2017, 38(9): 21-26. (in Chinese with English abstract)
DOI:10.7506/spkx1002-6630-201709004. http://www.spkx.net.cn
收稿日期:2016-05-06
基金项目:国家自然科学基金面上项目(31471604);国家农业科技创新工程项目;国家现代农业(肉羊)产业技术体系建设专项(CARS-39)
作者简介:高星(1990—),女,硕士研究生,研究方向为肉品科学与技术。E-mail:gaoxing123work@163.com
*通信作者:张德权(1972—),男,研究员,博士,研究方向为肉品科学与技术。E-mail:dequan_zhang0118@126.com