钟文秀,袁江兰*,李传雯,康 旭,何首春
(湖北工业大学生物工程与食品学院,湖北 武汉 430068)
摘 要:以高盐稀态法酿造米渣生酱油,探究菌种耦合对其品质和抗氧化活性的影响。4 组不同菌种耦合发酵的米渣生酱油品质测定结果表明其各项质量指标均达到GB 18186—2000《酿造酱油》中一级酱油的标准,其中S4生酱油(米曲霉耦合黑曲霉和鲁氏酵母发酵)游离氨基酸总量达到41.81 g/L,4 种米渣生酱油的总氨基酸、必需氨基酸、呈味氨基酸和抗氧化氨基酸的质量浓度均高于同时发酵的大豆生酱油S5生酱油(米曲霉发酵)。S3生酱油(米曲霉耦合鲁氏酵母发酵)和S4生酱油的抗氧化活性普遍高于S1生酱油(米曲霉发酵)和S2生酱油(米曲霉耦合黑曲霉发酵),各结果均表明S3的抗氧化活性最强,清除羟自由基的能力达到等质量浓度VC的最高175.42 倍,对1,1-二苯基-2-三硝基苯肼自由基清除能力达到VC的2.16 倍。抗氧化活性受发酵温度影响明显,分别在35 ℃和50 ℃发酵时出现峰值。多菌种耦合发酵对米渣生酱油品质和抗氧化活性均有明显改善,其中鲁氏酵母发挥了明显作用。
关键词:米渣生酱油;品质;抗氧化活性;菌种耦合;鲁氏酵母
米渣又常称为大米蛋白粉,为淀粉糖企业的大宗副产品,其蛋白质量分数达65%左右,是优质的植物蛋白资源。米渣蛋白极难溶解于水,导致其在食品领域的应用非常有限,而主要用作为饲料,未能充分实现其利用价值。米渣是酿造酱油的潜在优质原料 [1-3],以米渣完全替代大豆或豆粕,并辅以面粉、麸皮制曲和发酵可以制成品质良好的酱油。在酿造过程中微生物蛋白酶系不断将蛋白质降解成肽类、氨基酸等可溶性含氮物,加之细胞转化作用,使难溶于水的谷蛋白逐渐降解和转化而进入生酱油,从而使蛋白质利用率逐渐提高 [2]。目前,米渣生酱油蛋白质利用率可达70%。米渣生酱油不仅营养丰富,而且具有典型的酱油风味和独特的香气及滋味 [4],其价值始于米渣但又超越米渣。米渣生酱油不仅有利于丰富酱油品种,而且可以拓宽米渣的应用领域,从而大幅度提高米渣附加值。多菌种耦合发酵是发酵调味品的研究热点,李琴等 [5]采用双菌种混合制曲,促进了原料蛋白质与淀粉的降解,多菌种耦合发酵有利于酿造出优质酱油 [6]。课题组前期研究表明菌种耦合对米渣蛋白的利用率及产品品质具有重要影响,已经确定了菌种耦合发酵的方式和工艺,并且发现米渣生酱油具有明显的抗氧化活性 [2]。本实验在前期基础上进一步探究菌种耦合对米渣生酱油抗氧化活性和品质的影响。
1.1 材料与试剂
米渣 湖北德安府糖业有限责任公司;米曲霉HN3.042、黑曲霉AS3.350 上海佳民酿造食品有限公司酿造一厂;鲁氏酵母 安琪酵母股份有限公司;茚三酮显示液 日本日立公司;2,2’-联氮-二(3-乙基-苯并噻唑-6-磺酸)二铵盐(2,2’-azinobis-(3-ethylbenzthiazoline-6-sulphonate,ABTS)、罗丹明6G(高级纯) 阿拉丁试剂(上海)有限公司;1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH) 上海源叶生物科技有限公司。
1.2 仪器与设备
L-8900全自动氨基酸分析仪、F-7000荧光分光光度计日本日立公司;JK9870A自动凯氏定氮仪 济南海能仪器股份有限公司;AAnalyst400原子吸收光谱仪 珀金埃尔默仪器(上海)有限公司;Unico 2100可见分光光度计 尤尼柯(上海)仪器有限公司。
1.3 方法
1.3.1 米渣生酱油制备
按照m(米渣)∶m(麸皮)∶m(面粉)∶V(水)=6.0∶2.0∶2.0∶7.8的配比将原料混合均匀,然后蒸料并冷却至35 ℃以下,接种、制曲、发酵而成。其中,制曲条件为:曲料接种后在温度32~35 ℃、相对湿度85%~90%的恒温恒湿曲室中培养36 h,得成曲。发酵条件为:成曲装入1 L可密封罐中,添加2.5 倍体积盐水混合制成质量分数为10.86%盐的稀醪,30~35 ℃条件下恒温发酵4 个月,过滤得生酱油。
共5 个样品,分别标记为S1、S2、S3、S4、S5,其中S1为接种质量分数0.2%米曲霉进行制曲并发酵而得的生酱油;S2为接种0.16%米曲霉和0.04%黑曲霉混菌制曲并发酵而得的生酱油;S3为接种0.2%米曲霉制曲并在发酵初始接种0.05%鲁氏酵母混菌发酵而得的生酱油,S4为接种0.16%米曲霉和0.04%黑曲霉混菌制曲并在发酵初始接种0.05%鲁氏酵母混菌发酵而得的生酱油,S5为大豆生酱油,只以大豆取代米渣,其他条件均同S1。制备方法、菌种耦合方式和接种量均参考前期结果 [4]。
1.3.2 基本理化指标分析
总氮、总固形物及食盐含量按照GB 18 186—2000《酿造酱油》 [7]标准方法检测。无盐固形物计算方式为总固形物减去盐含量。总酸根据GB/T 12456—2008《食品中总酸的测定》 [8]测定。还原糖含量根据GB 5009.7—2008《食品中还原糖的测定》 [9]测定。γ-氨基丁酸测定方法与游离氨基酸检测方法(1.3.3.2节)相同。
1.3.3 酱油成分及抗氧化活性检测
1.3.3.1 微量元素检测
根据文献[10]中原子吸收光谱法测定。
1.3.3.2 氨基酸分析
取适量酱油样品与等体积的10%磺基水杨酸溶液,混合均匀后于4 ℃条件下放置15 min,15 00 0 r/min离心15 min,取上清液用0.02 mol/L盐酸稀释200 倍,用0.22 μm微孔滤头过滤后采用氨基酸自动分析仪检测。
样品经离子交换柱(4 0 m m×6 0 m m)以0.40 mL/min流速洗脱后,经反应柱由茚三酮及茚三酮缓冲溶液于流速0.35 mL/min、柱温135 ℃、柱压0.982 MPa条件下反应,样品分析周期为53 min。
1.3.3.3 还原力检测
采用普鲁士蓝法 [11]测定还原力。酸性条件下具抗氧化活性的物质将铁氰化钾(K 3Fe(CN) 6)还原成亚铁氰化钾(K 4Fe(CN) 6),然后还原产物与Fe 3+生成普鲁士蓝,其A 700 nm与还原力呈正比。以100 μg/mL VC溶液的还原力标准曲线表示每毫升酱油的还原力与VC溶液质量浓度的等同量,即μg VC/mL(μg AAE/mL)。
1.3.3.4 DPPH自由基清除能力
参考Ohnish等 [12]的方法。DPPH自由基清除率按公式(1)计算。DPPH-乙醇溶液浓度为0.15 mmol/L,样品稀释200 倍进行测定并以抗坏血酸溶液为阳性对照。
式中:A 样品为等体积样品稀释液和DPPH乙醇溶液的吸光度;A 对照为等体积样品稀释液和无水乙醇的吸光度;A 空白为等体积水和DPPH乙醇溶液的吸光度。
测定5 个浓度下样品对DPPH自由基清除能力,经线性回归后计算出DPPH自由基清除率为50%时的酱油质量浓度,即EC 50值/(μg/mL)。
1.3.3.5 总抗氧化能力检测
采用ABTS法 [13]测定总抗氧化能力。各取5 0 m L 2 mmol/L ABTS溶液与70 mmol/L K 2S 2O 8溶液混合均匀,室温避光放置12~16 h,得ABTS储备液。用磷酸盐缓冲溶液(phosphate buffer saline,PBS)(10 mmol/L,p H 7.4)将A B T S储备液稀释,使其A 7 3 4 n m为0.700±0.020。取0.2 mL样品稀释液与3.8 mL ABTS工作液混合均匀反应4 min后检测其A 734 nm,ABTS +·清除能力与A 734 nm呈反比。以Trolox为阳性对照,并绘制标准曲线,样品的抗氧化能力用当量抗氧化能力(Troloxequivalent antioxidant capacity,TEAC)表示。
1.3.3.6 羟自由基(·OH)清除能力检测
采用罗丹明6G-Fenton体系荧光法 [14]。在10 mL具塞比色管中,依次加入0.5 mL pH 4.8 PBS、0.5 mL 0.1 mg/L罗丹明6 G溶液、1.0 m L 1.0 mmo l/L Fe SO 4溶液、0.6 m L 0.0 3% H 2O 2溶液,加水至刻度,混匀,反应15 min。λ ex、λ em分别为348、550 nm,激发和发射狭缝宽度为5 nm,以不加Fenton试剂的体系为空白,其荧光强度为F 0,0.3 mL样品稀释液与罗丹明6G-Fenton溶液体系的荧光强度记为F samp le,并以0.53 mg/mL抗坏血酸溶液为阳性对照。·OH清除率按公式(2)计算。
式中:F sample-F为样品清除罗丹明6G-Fenton体系·OH的量;F 0-F为罗丹明6G-Fenton体系所产生的·OH的量。
测定5 个质量浓度下样品对·OH清除能力,经线性回归后计算EC 50值(concentration for 50% of maximal effect,EC 50),即·OH清除率为50%时的酱油质量浓度,即EC 50值/(μg/mL)。
1.3.4 感官质量评价
根据GB 18186—2000对不同级别酿造酱油感官质量的评价 [7],制定感官评分表(表1)。组织10 位食品专业人员(男女各半)对米渣生酱油进行感官评定,为保证感官评定的准确性,取样适量,样品随机编号、呈样,结果取3 次评分的平均值。
表1 感官综合评价
Table 1 Criteria for sensory evaluation of rice sauce
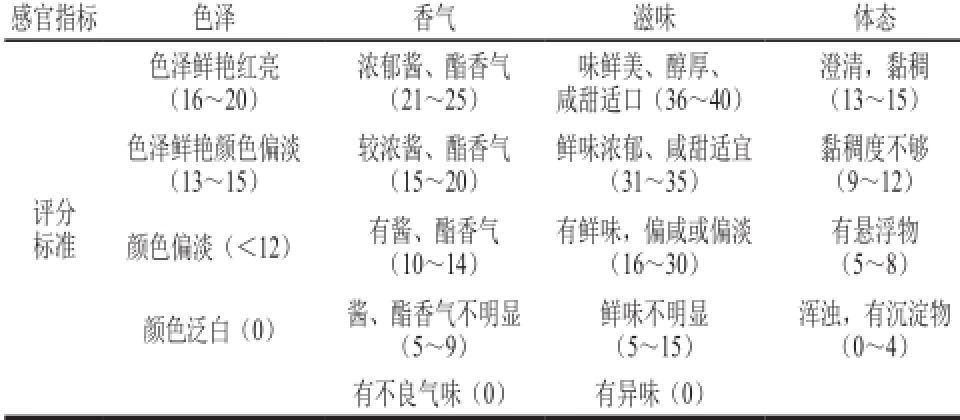
感官指标色泽香气滋味体态澄清,黏稠(13~15)色泽鲜艳颜色偏淡(13~15)色泽鲜艳红亮(16~20)浓郁酱、酯香气(21~25)味鲜美、醇厚、咸甜适口(36~40)黏稠度不够(9~12)颜色偏淡(<12)有酱、酯香气(10~14)较浓酱、酯香气(15~20)鲜味浓郁、咸甜适宜(31~35)评分标准有悬浮物(5~8)颜色泛白(0)酱、酯香气不明显(5~9)有鲜味,偏咸或偏淡(16~30)浑浊,有沉淀物(0~4)有不良气味(0)有异味(0)鲜味不明显(5~15)
1.3.5 生酱油的体外抗氧化活性比较
S1~S5在35 ℃条件下恒温发酵4 个月。以还原力、DPPH自由基清除能力、总抗氧化能力和·OH清除能力为指标评价其体外抗氧化活性的大小。
1.3.6 发酵温度对米渣生酱油抗氧化活性和品质的影响
S1~S4在30、35、40、45、50 ℃ 5 个温度水平下发酵4 个月。以氨基态氮、全氮、总还原力、DPPH自由基清除能力为指标评价发酵温度对生酱油抗氧化活性和品质的影响。
1.3.7 发酵时间对米渣生酱油抗氧化活性和品质的影响
S1~S4在35 ℃恒温发酵,分别于1、3、5、7、10、40、70、90、120 d取样,测定还原糖、氨基态氮、总还原力、DPPH自由基清除能力,评价发酵过程中样品抗氧化能力和品质的动态变化规律。
1.4 数据统计与分析
每个样品均做6 个平行(即6 批发酵样品),结果以±s表示。所有数据利用Origin 8.0统计和绘图,SPSS 18.0软件进行差异性分析,同一系列不同字母表示差异显著(P<0.05)。
2.1 米渣生酱油的基本理化指标由表2可知,相同条件下所制得的米渣生酱油和大豆生酱油各项指标均到达国家一级酱油标准,其中氨基态氮含量远高于特级酱油标准(0.8 g/100 mL)。以添加黑曲霉发酵的S2和S4酱油中的氨基态氮含量最大,更接近大豆生酱油S5,可能是黑曲霉和米曲霉的蛋白酶系互补和协同作用的结果,提高了米渣蛋白的降解与转化率。黑曲霉产酸性蛋白酶和淀粉酶的能力较强,有利于提高酱油中氨基态氮、总氨基酸、谷氨酸及还原糖的含量 [15]。酱油中微量元素的含量均达到国标,铅含量低于
表2 米渣生酱油基本理化指标和微量元素
Table 2 Nutritional components and trace elements of rice sauce g/100 mL
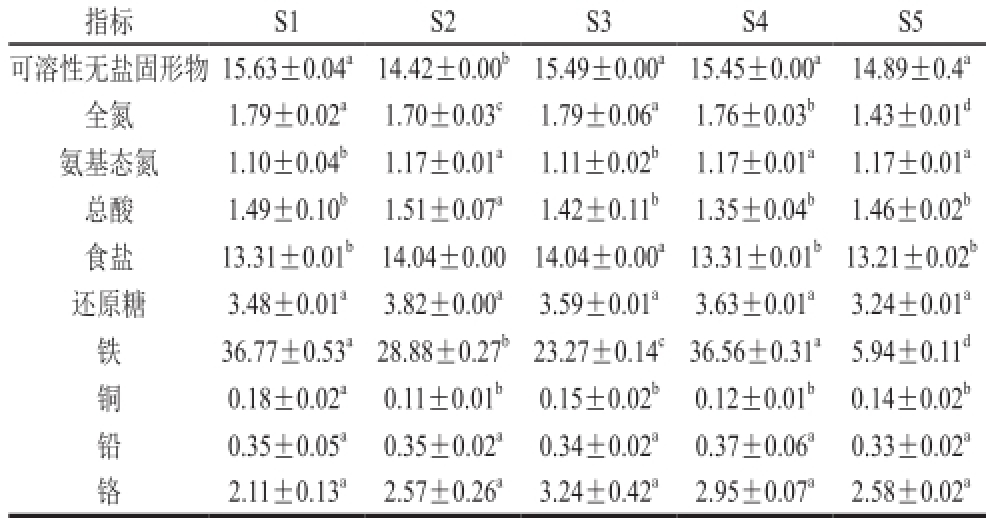
注:同行小写字母不同表示差异显著(P<0.05)。表3~5同。
指标S1 S2 S3 S4 S5可溶性无盐固形物15.63±0.04 a14.42±0.00 b15.49±0.00 a15.45±0.00 a14.89±0.4 a全氮1.79±0.02 a1.70±0.03 c1.79±0.06 a1.76±0.03 b1.43±0.01 d氨基态氮1.10±0.04 b1.17±0.01 a1.11±0.02 b1.17±0.01 a1.17±0.01 a总酸1.49±0.10 b1.51±0.07 a1.42±0.11 b1.35±0.04 b1.46±0.02 b食盐13.31±0.01 b14.04±0.00 14.04±0.00 a13.31±0.01 b13.21±0.02 b还原糖3.48±0.01 a3.82±0.00 a3.59±0.01 a3.63±0.01 a3.24±0.01 a铁36.77±0.53 a28.88±0.27 b23.27±0.14 c36.56±0.31 a5.94±0.11 d铜0.18±0.02 a0.11±0.01 b0.15±0.02 b0.12±0.01 b0.14±0.02 b铅0.35±0.05 a0.35±0.02 a0.34±0.02 a0.37±0.06 a0.33±0.02 a铬2.11±0.13 a2.57±0.26 a3.24±0.42 a2.95±0.07 a2.58±0.02 a
1 mg/kg,符合国家卫生标准。
2.2 米渣生酱油的游离氨基酸
表3 生酱油游离氨基酸组成
Table 3 Free amino acid composition of rice sauce

注:A.鲜味氨基酸;B.人体必需氨基酸;C.抗氧化氨基酸;TAA.总氨基酸总量;DTAA.鲜味氨基酸总量;EAA.必需氨基酸总量;FAA.呈味氨基酸;AAA.抗氧化氨基酸总量。
氨基酸S1 S2 S3 S4 S5天冬氨酸 A/(g/L)3.26±0.01 b3.41±0.02 b3.11±0.00 b3.70±0.01 a3.73±0.03 a苏氨酸 B/(g/L)1.69±0.02 a1.78±0.02 a1.61±0.00 a1.88±0.00 a1.61±0.02 a丝氨酸/(g/L)2.19±0.03 a2.29±0.02 a2.10±0.00 a2.45±0.01 a2.13±0.01 a谷氨酸 A/(g/L)3.73±0.07 e4.09±0.01 c5.10±0.07 b5.84±0.01 a4.43±0.02 d甘氨酸 A/(g/L)1.59±0.03 a1.68±0.01 a1.53±0.03 a1.72±0.01 a1.33±0.00 b丙氨酸 A/(g/L)2.34±0.03 a2.45±0.01 a2.21±0.02 a2.47±0.02 a1.83±0.01 b半胱氨酸 C/(g/L)0.39±0.01 a0.40±0.01 a0.38±0.00 a0.40±0.01 a0.35±0.01 a缬氨酸 B/(g/L)3.01±0.03 a3.12±0.02 a2.93±0.04 a3.21±0.01 a2.46±0.03 b甲硫氨酸 BC/(g/L)1.28±0.04 a1.30±0.01 a1.42±0.01 a1.31±0.02 a0.99±0.01 b异亮氨酸 B/(g/L)3.01±0.01 b3.15±0.00 b2.87±0.01 b3.41±0.07 a3.44±0.03 a亮氨酸 B/(g/L)3.09±0.01 a3.22±0.01 a3.01±0.05 a3.31±0.04 a2.84±0.01 b酪氨酸 AC/(g/L)1.54±0.02 a1.56±0.00 a1.58±0.02 a1.63±0.00 a1.54±0.00 a苯丙氨酸 AB/(g/L)2.19±0.01 a2.26±0.01 a2.20±0.00 a2.34±0.01 a2.01±0.01 a赖氨酸 B/(g/L)1.79±0.00 b1.90±0.02 b1.73±0.03 b1.84±0.02 b2.36±0.01 a组氨酸 C/(g/L)1.02±0.02 a1.06±0.01 a1.01±0.00 a1.08±0.01 a0.99±0.01 a精氨酸 B/(g/L)3.05±0.01 b3.14±0.03 b2.93±0.06 b3.38±0.02 a2.51±0.02 c脯氨酸/(g/L)1.27±0.01 a1.43±0.00 a1.29±0.01 a1.65±0.01 a1.16±0.01 aγ-氨基丁酸/(g/L)0.24±0.01 a0.26±0.01 a0.24±0.00 a0.21±0.00 a0.10±0.01 b总氨基酸含量/(g/L)36.70±0.05 b38.53±0.06 b37.27±0.03 b41.81±0.04 a35.92±0.03 b鲜味氨基酸/(g/L)14.66±0.01 c15.46±0.02 b15.73±0.01 b17.69±0.01 a14.91±0.01 c必需氨基酸/(g/L)16.07±0.01 c16.74±0.01 b15.78±0.01 c17.30±0.02 a15.74±0.01 c呈味氨基酸/(g/L)19.80±0.01 c20.97±0.03 b20.74±0.02 b23.67±0.02 a19.81±0.01 c抗氧化氨基酸/(g/L)4.24±0.01 a4.34±0.023 a4.40±0.01 a4.42±0.03 a3.88±0.02 bDTAA/TAA/%39.94±0.05 a40.12±0.01 a42.21±0.01 a42.31±0.01 a41.51±0.00 aEAA/TAA/%43.80±0.01 a43.44±0.02 a42.33±0.02 b41.37±0.03 b43.82±0.01 aFAA/TAA/%53.97±0.03 c54.41±0.02 c55.66±0.02 b56.60±0.02 a55.12±0.03 bAAA/TAA/%11.56±0.00 a11.25±0.01 a11.81±0.01 a10.58±0.02 a10.77±0.02 a
由表3可知,米渣生酱油氨基酸总量高于传统大豆生酱油(P<0.05),4 种米渣生酱油总氨基酸含量大小排序依次为:S4>S2>S3>S1,其中S4比S1提高了13.94%(P<0.05),S2比S1提高了5.00%(P<0.05),而S3比S1仅提高1.55%(P>0.05),说明多菌种耦合由于蛋白酶系丰富而更有利于蛋白质降解和氨基酸形成,其中发挥主要作用的是米曲霉,而黑曲霉蛋白酶活力明显低于米曲霉,故贡献次于米曲霉,酵母菌贡献最小。酱油含丰富的游离氨基酸,但各氨基酸质量浓度差别较大,S4中天冬氨酸、谷氨酸、缬氨酸、亮氨酸、异亮氨酸和精氨酸6 种氨基酸质量浓度均高于其他3 种米渣生酱油,其中谷氨酸和天冬氨酸质量浓度分别为5.84 g/L和3.70 g/L,分别是S1酱油的1.56、1.13 倍,S2酱油的1.43、1.11 倍,S3的1.14、1.19 倍(P<0.05),因此4 种米渣生酱油鲜味氨基酸质量浓度顺序依次为S4>S3>S2>S1。采用高盐稀态法酿造酱油往往会由于酱油含盐量偏大而导致咸味过重,但米渣生酱油中呈味氨基酸质量浓度高,所以呈味协调性很好,具有咸甜适宜、鲜味浓、总体滋味醇厚持久等特点。4 种米渣生酱油呈味氨基酸质量浓度顺序依次为S4>S3>S2>S1。4 种米渣生酱油必需氨基酸总量(除色氨酸未检测外)所占总氨基酸比例均高于40%,符合FAO/WHO理想蛋白质标准。米渣酱油中除含丰富的必需氨基酸和呈味氨基酸外,还含有较为丰富的γ-氨基丁酸,其质量浓度高于200 mg/L。三菌耦合发酵所得生酱油总氨基酸含量及谷氨酸含量较单一米曲霉发酵的S1酱油均明显增加,对提高生酱油的感官质量非常有利。多菌种耦合制曲酿造大豆生酱油也有相似结果 [16-18]。
2.3 米渣生酱油的感官质量
表4 米渣生酱油的感官质量评分
Table 4 Sensory evaluation scores of rice sauce

感官指标S1 S2 S3 S4 S5色泽(0~20)18.00±0.43 c17.00±0.63 d20.11±0.56 a19.00±0.45 b16.00±0.70 e香气(0~25)18.00±0.45 c20.00±0.45 b21.00±1.14 b23.00±0.78 a24.00±0.37 a滋味(0~40)28.00±0.45 d31.23±1.84 c35.11±0.56 b38.00±0.45 a39.00±0.51 a体态(0~15)15.00±0.00 a15.00±0.00 a15.00±0.00 a15.00±0.00 a15.00±0.00 a总分(100)79.00±0.23 d83.00±1.21 c91.00±0.45 b95.00±0.78 a95.00±0.57 a
由表4可知,由耦合菌种发酵的米渣生酱油感官质量总体上优于单一菌种发酵的产品,与文献研究结果一致 [4]。多菌种耦合发酵有利于产生丰富酶系,提高原料利用率及丰富酱油组成。发酵4 个月的米渣生酱油呈色优于大豆生酱油,这可能与原料本身化学组成有关,三菌耦合发酵的米渣生酱油S4感官质量与大豆生酱油相近。米渣生酱油呈红褐色、有光泽、透明度好、无沉淀,具有很强的酱香味和酯香味,味道鲜美、醇厚、咸甜适宜、后味浓厚,达到GB 18186—2000高盐稀态酱油一级酱油的标准,其中S3和S4酱油感官质量均明显好于S1和S2(P<0.05),这可能与酵母参与发酵有关。现有研究表明,耐盐酵母是提高酱油香气的关键,添加耐盐酵母协同制曲和发酵显著增加了酱油风味物质的种类和典型风味物质的含量 [19-20]。其中鲁氏酵母的添加,增加了主发酵过程中醇类、糖醇类及呋喃酮类物质的生成并在后期产生呈鲜味的核酸类物质 [21],赋予酱油醇厚的滋味和愉悦的香气。
2.4 米渣生酱油的抗氧化活性
由图1可知,4 种米渣生酱油的DPPH自由基清除能力、总抗氧化能力及·OH清除能力均优于大豆生酱油。单一米曲霉发酵时,同样条件下,大豆生酱油的还原力略强于米渣生酱油,而DPPH自由基清除能力、总抗氧化能力和·OH清除能力均比米渣生酱油弱,多菌种耦合发酵对抗氧化能力的提高有重要贡献。由图1a可知,米渣生酱油的还原力与发酵菌种密切相关,S3还原力最强,S4次之,S1最弱(P<0.05),可能酵母参与发酵对还原力有较大贡献。由图1b可知,稀释200 倍的4 种酱油的DPPH自由基清除率为57.53%~68.61%,均高于与5 μg/mL VC溶液的DPPH自由基清除率(P<0.05),其中S3最高,S4、S1次之,S2最弱。由图1c可知,4 种米渣生酱油总抗氧化能力TEAC值均大于50,S3依然最高(P<0.05)。由图1d可知,稀释200 倍的4 种米渣生酱油对·OH清除率为68.41%~82.26%(P<0.05),高于0.53 mg/mL VC溶液,顺序依次为S3、S4、S2、S1。菌种耦合发酵对自由基清除能力明显高于单一米曲霉发酵。·OH是人体正常新陈代谢过程中产生的具有代表性的自由基,是目前已知活性氧中对生物体毒性最强、危害最大、氧化能力极强的一种自由基,它会使细胞内的糖、蛋白质、核酸及脂类等物质发生氧化反应,进而造成组织过氧化及细胞膜的损伤 [22],米渣生酱油可能对人体有潜在的抗氧化保护作用,综合分析表明S3和S4抗氧化活性均较强,说明添加酵母对酱油抗氧化活性有显著影响。有研究表明混合酵母菌发酵红茶菌也明显提高了红茶菌的抗氧化活性 [23],与本结果一致表明酵母参与发酵对抗氧化活性贡献明显。


图1 米渣生酱油的抗氧化活性
Fig. 1 Antioxidant activities of rice sauce
表5 4 种米渣生酱油与VC溶液的EC
50
Table 5 EC
50values of four rice sauces and VC for scavenging of DPPH and hydroxyl radicals
μg/mL

样品DPPH自由基·OH S1 3.41±0.03 b3.19±0.11 bS2 3.31±0.01 b2.87±0.05 bS3 2.96±0.04 b2.51±0.02 bS4 3.23±0.01 b2.57±0.01 bVC 6.39±0.01 a440.30±0.78 a
各样品EC 50如表5所示,米渣生酱油抗氧化能力明显优于VC溶液,4 种生酱油对DPPH自由基清除能力分别约是VC溶液的1.87、1.93、2.16、1.98 倍(P<0.05),以S3酱油最佳。4 种生酱油均具有很强的清除·OH能力,其清除能力分别是VC溶液的138.02、153.4、175.42、171.32 倍(P<0.05),表明菌种耦合发酵对自由基清除能力明显高于单一菌种发酵。鲁氏酵母协同发酵在丰富产品风味的同时也在一定程度提高了产品的抗氧化活性。鲁氏酵母除了可分泌出高浓度的多元醇外,还具有中强抗氧化活性,可作为天然抗氧化剂的来源 [24]。
2.5 发酵温度对生酱油抗氧化活性和品质的影响
由图2可知,发酵温度对米渣生酱油氨基态氮、全氮及抗氧化活性的影响明显。氨基态氮和总氮含量在30~50 ℃条件下时,随发酵温度的升高先增大后逐渐减小,并在35 ℃时达到最大值,各酱油氨基态氮和全氮含量分别高于1.1 g/100 mL和1.7 g/100 mL(P<0.05),由于微生物具有适宜的环境生长范围,其分泌的酶系统的活力受温度影响进而直接影响原料的降解及其物质的生成,同时也反映了中低温发酵酱油有利于提高其氨基态氮的生成率,这可能与低温有利于保持酶活力相关 [25]。发酵温度为30 ℃的酱油中氨基态氮和总氮含量略低于35 ℃(P<0.05),可能也与发酵周期相关。当发酵温度达5 0 ℃时,各酱油的氨基态氮和全氮含量均为最低值,高温加速了氨基酸和糖类之间美拉德反应的产生,消耗了氨基酸 [24]。当发酵温度为30~45 ℃时,生酱油还原力和DPPH自由基清除率先随发酵温度的升高而增大,在发酵温度为35 ℃时4 种米渣生酱油还原力和DPPH自由基清除率均达到最大值,然后随温度升高而明显降低(P<0.05)。50 ℃条件下发酵的生酱油还原力和DPPH自由基清除率均陡增(P<0.05),可能是由于高温促进了美拉德反应,使类黑精等产物快速累积,从而提高了生酱油的抗氧化能力 [26]。各发酵温度酿造酱油的还原力和DPPH自由基清除能力均以S3为最大(P<0.05),进一步说明酵母在发酵过程中对抗氧化的突出贡献。米渣生酱油品质和抗氧化活性均与发酵温度密切相关,温度影响了发酵体系酶的产生及活力、菌相和美拉德反应速度,从而导致酱油滋味、香气、色泽和抗氧化成分的差异 [2 7-2 8]。
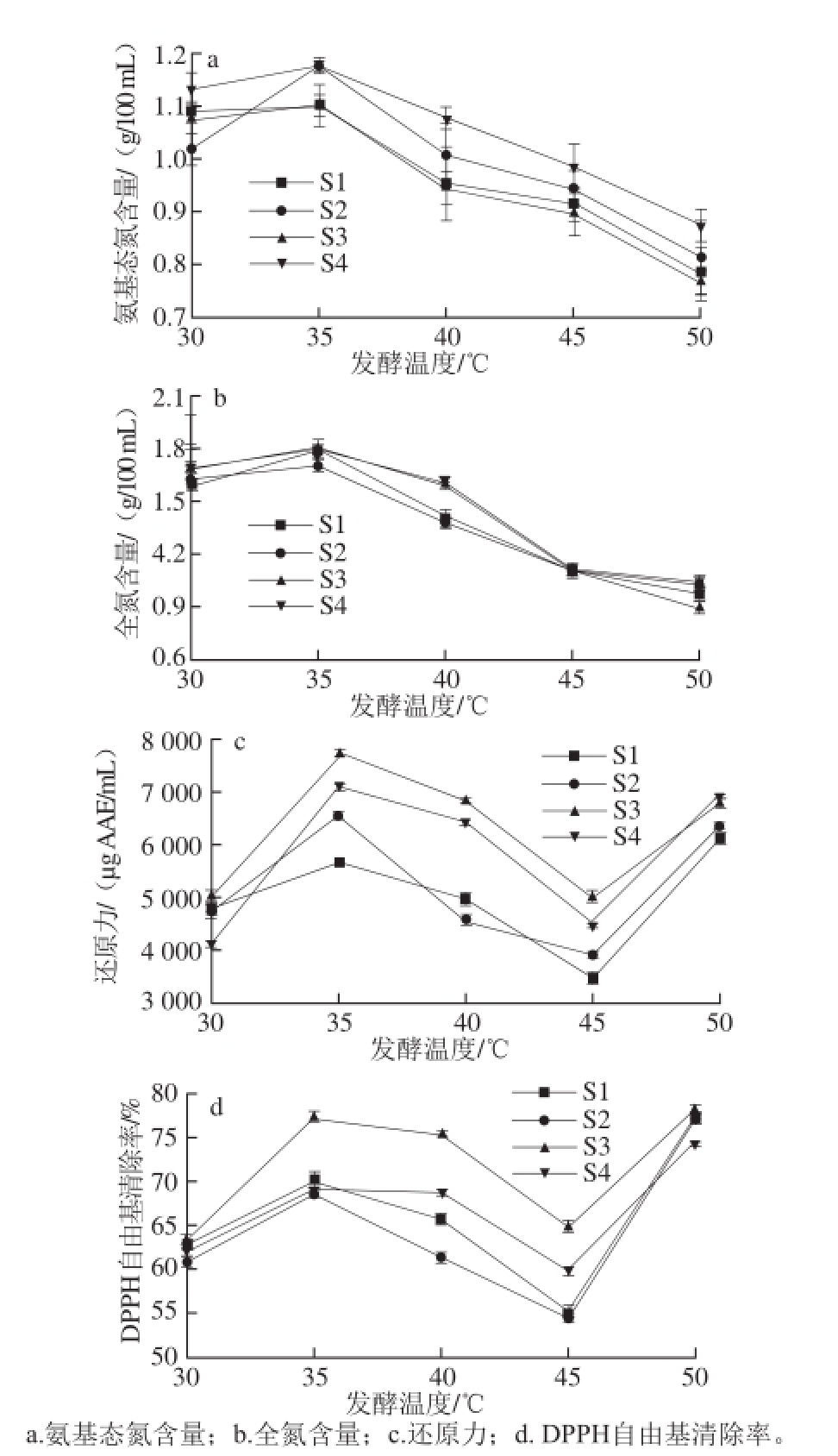
图2 发酵温度对米渣生酱油抗氧化的影响
Fig. 2 Effect of fermentation temperature on antioxidant activity of rice sauce
2.6 发酵时间对生酱油还原糖含量、氨基态氮及抗氧化活性的影响
如图3a所示,S1~S4还原糖含量均随发酵时间的延长呈现先增加后缓慢减少再趋于稳定的趋势,可能与美拉德反应消耗了还原糖有关,在发酵成熟期,还原糖含量趋于稳定 [28]。酱油氨基态氮(图3b)含量随发酵时间的延长而增加,发酵前期由于蛋白酶的水解作用,氨基态氮含量增长较快,发酵中后期氨基态氮含量增长速度减缓并于发酵成熟期趋于稳定,其中S2和S4的氨基态氮含量明显高于S1和S3(P<0.05),说明黑曲霉可能对氮素的转化有明显作用。各生酱油的还原力随发酵时间的延长而增大(图3c)。在发酵前期,发酵液还原能力增幅大(P<0.05),发酵中后期,还原力增速变缓,可能与酱油发酵成熟其各组分趋于稳定有关,在整个发酵期间,S3的还原力始终最高,S4次之。4 种生酱油对DPPH自由基的清除能力(图3d)随发酵时间的变化趋势相同,随发酵时间的延长,酱油对DPPH自由基的清除能力由弱变强。与等质量浓度的VC(5 μg/mL)相比,前3 d发酵的酱油发酵液对DPPH自由基的清除能力均低于VC溶液,发酵5 d后,S1的DPPH自由基清除率高于S2、S3、S4和VC溶液(P<0.05)。在酱油发酵前期,单菌种发酵在初始5 d内分解率较高,在发酵5~10 d内,单菌种酶解作用弱于耦合菌种,因此耦合菌种发酵的生酱油DPPH自由基清除能力增幅大于单菌种发酵酱油(P<0.05)。发酵10 d后,4 种生酱油样品对DPPH自由基的清除能力均明显大于VC溶液(P<0.05)。发酵中后期,酱油中抗氧化活性成分逐渐趋于稳定,对DPPH自由基的清除能力也趋于稳定(P>0.05)。
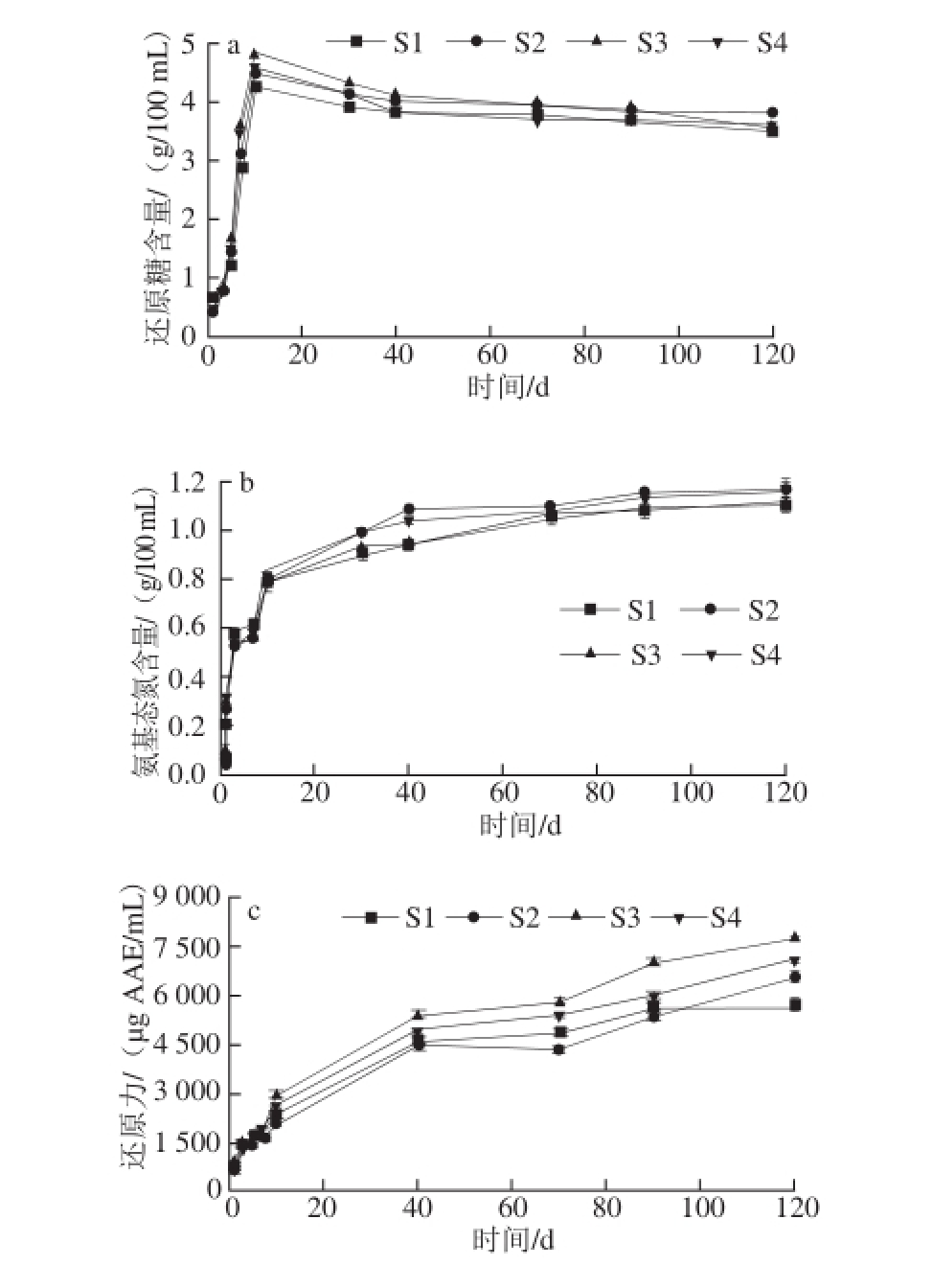
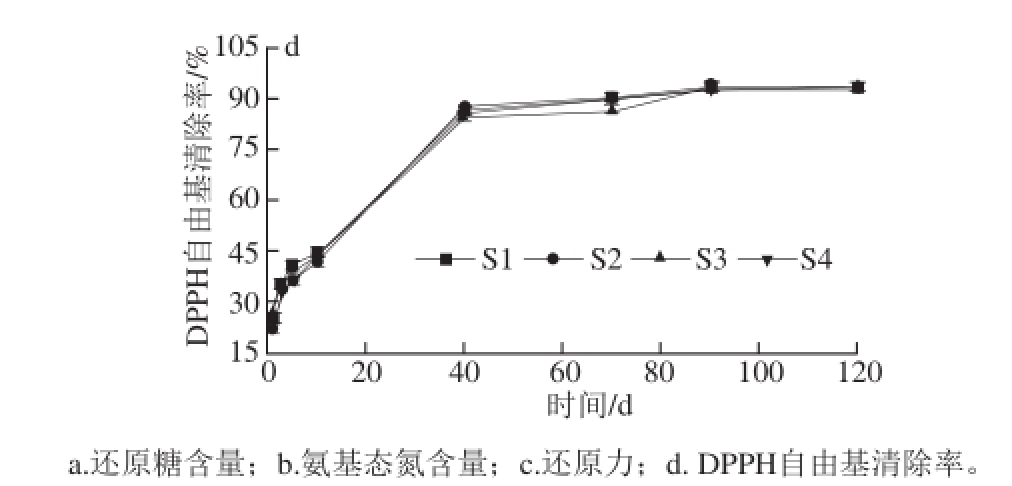
图3 发酵时间与米渣生酱油抗氧化活性
Fig. 3 Correlation between fermentation time and antioxidant activity of rice sauce
以米曲霉为主要菌种,耦合黑曲霉和酵母酿造的米渣生酱油,其感官品质、氨基态氮含量、氨基酸组成和总量均高于单一菌种发酵的米渣生酱油,并以三菌种耦合发酵的S4生酱油品质最佳,米曲霉耦合酵母发酵的S3略低于S4,但S3、S4的抗氧化能力均较强。温度对米渣生酱油抗氧化活性的影响显著,35 ℃条件下恒温发酵的米渣生酱油抗氧化活性高,但是50 ℃条件下发酵时各样品的抗氧化活性均出现新的峰值。随发酵时间延长,各米渣生酱油抗氧化活性增加,特别是在发酵初始的10 d内增加迅速,而发酵成熟期,抗氧化活性也趋于平稳。不同菌种发酵的米渣生酱油表现出不同的抗氧化活性,4 种米渣生酱油清除DPPH自由基和·OH效果均优于VC,以S3、S4清除效果较好,暗示酵母对抗氧化活性的提高有明显作用。
参考文献:
[1] 靳艳, 袁江兰, 李文竹, 等. 米渣生酱油生产工艺优化[J]. 食品科技, 2013, 38(4): 290-293. DOI:10.13684/j.cnki.spkj.2013.04.033.
[2] 林灿, 袁江兰, 邓冲, 等. 米渣蛋白酱油发酵中氮素演变规律与过程调控相关性[J]. 现代食品科技, 2015, 31(6): 116-121. DOI:10.13982/ j.mfst.1673-9078.2015.6.019.
[3] YUAN J L, ZHONG W X, JIN Y, et al. Fermentation technology and quality of sauce prepared from rice dreg protein[J]. Journal of the Institute of Brewing, 2015, 121: 449-454. DOI:10.1002/jib.241.
[4] 邓冲, 钟文秀, 李传雯, 等. 米渣酱油与传统大豆酱油酿造过程的动态研究[J]. 中国调味品, 2016, 41(4): 1-6. DOI:10.3969/ j.issn.1000-9973.2016.04.001.
[5] 李琴, 杜风刚. 双菌种制曲在酱油生产中的应用[J]. 中国调味品, 2003, 28(12): 36-38. DOI:10.3969/j.issn.1000-9973.2003.12.011.
[6] 张海珍, 蒋予箭, 陈敏. 多菌种制曲与发酵在酿造酱油中的应用现状[J]. 中国酿造, 2 0 08, 27(9): 1-4. DOI:1 0.3 969/ j.issn.0254-5071.2008.09.001.
[7] 张林, 鲁肇元, 李栓勤, 等. 酿造酱油: GB 18186—2000[S]. 北京:中国标准出版社, 2000: 3-6.
[8] 杨大进, 常迪, 赵馨,等. 食品中还原糖的测定: GB 5009.7—2008[S].北京: 中国标准出版社, 2009: 1-3.
[9] 龚玲娣, 徐清渠. 食品中总酸的测定: GB/T 12456—2008[S]. 北京:中国标准出版社, 2009: 1-5.
[10] 耿薇, 张君才, 郑敏燕, 等. 原子吸收光谱法检测饮用矿物质水中的金属元素[J]. 应用化工, 2010, 39(4): 600-601. DOI:10.3969/ j.issn.1671-3206.2010.04.037.
[11] OYAIZU M. Studies on products of browning reaction: antioxidant activities of products of browning reaction prepared from glucose amine[J]. Japanese Journal of Nutrition, 1986, 44(6): 307-315. DOI:10.5264/eiyogakuzashi.44.307.
[12] OHNISHI M, MORISHITA H, IWAHASHI H, et al. Inhibitory effects of chlorogenic acids on linoleic acid peroxidation and heamolysis[J]. Phytochemistry, 1994, 36(3): 579-583. DOI:10.1016/S 0031-9422(00)89778-2.
[13] RE R, PELLEGRINI N, PROTEGGERE A, A. et al. Antioxidant activity applying improved ABTS radical cation decolorization assay[J]. Free Radical Biology and Medicine, 1999, 26(8): 1231-1237. DOI:0.1016/S0891-5849(98)00315-3.
[14] 岳保旺, 范洪毓, 武宇芳, 等. 罗丹明6G-Fenton体系荧光法测定中草药抗氧化活性[J]. 分析科学学报, 2013, 29(5): 723-725.
[15] GAO X L, ZHAO H F, FENG Y Z, et al. A comparative study on physicochemical properties of Chinese-type soy sauces prepared using pure koji and mixed kojis[J]. African Journal of Biotechnology, 2010, 9(40): 6740-6747. DOI:10.5897/AJB10.767.
[16] ISHIHARA K, HONMA N, MATSUMOTO I, et al. Comparison of volatile components in soy sauce (Koikuchi Shoyu) produced using Aspergillus sojae and Aspergillus oryzae[J]. Nippon Shokuhin Kagaku Kogaku Kaishi, 1996, 43(9): 1063-1074. DOI:10.3136/nskkk.43.1063.
[17] UEKI T, NODA Y, TERAMOTO Y, et al. Practical soy sauce produ ctio n using a mixed Ko ji-making system[J]. Jo urnal of Fermen ta tion & Bioen g in eering, 199 4, 7 8(3): 262-264. DOI:10.1016/0922-338X(94)90302-6.
[18] 李保英. 多菌种酱油制曲工艺及其对酱油风味影响的研究[D].杭州: 浙江工商大学, 2013: 50-69.
[19] 刘晶晶, 胡勇, 陈梦, 等. 酱油酿造中米曲霉与风味酵母协同制曲的研究[J]. 现代食品科技, 2015, 31(9): 230-237. DOI:10.13982/ j.mfst.1673-9078.2015.9.038.
[20] 夏强, 刘奕雯. 耐盐酵母菌对发酵酱油风味作用及应用研究[J].商品与质量(学术观察), 2015(2): 239-239. DOI:10.3969/j.issn.1006-656X(x).2015.02.231.
[21] 徐莹, 姜维, 何晓霞. 耐盐性鲁氏酵母的研究进展[J]. 中国酿造, 2009, 28(10): 1-4. DOI:10.3969/j.issn.0254-5071.2009.10.001.
[22] 李勇, 孔令青, 高洪, 等. 自由基与疾病研究进展[J]. 动物医学进展, 2008, 29(4): 85-88. DOI:10.3969/j.issn.1007-5038.2008.04.021.
[24] MALBASA R V, LONCAR E S, VITAS J S, et al. Influence of starter cultures on the antioxidant activity of kombucha beverage[J]. Fo od Che mistry, 20 11, 127(4): 17 27-17 31. DOI:10.101 6/ j.foodchem.2011.02.048.
[25] NAYLIN N, TAING O, HASHINAGA F, et a l. An tioxidant activity of sugar-tolerant yeast Zyg osaccharomyces rouxii[J]. Food Biotechnology, 2005, 19(2): 107-120. DOI:10.1081/FBT-200063435.
[26] HOFMANN T. Studies on the relationship between molecular weight and the color potency of fractions obtained by thermal treatment of glucose/amino acid and glucose/protein solutions by using ultracentrifugation and color dilution techniques[J]. Journal of Agricultural and Food Chemistry, 1998, 46(10): 3891-3895. DOI:0.1021/jf980397e.
[27] NAKAHARA T, YAMAGUCHI H, UCHIDA R. Effect of temperature on the stability of various peptidases during peptide-enriched soy sauce fermentation[J]. Journal of Bioscience and Bioengineering, 2011, 113(3): 355-359. DOI:10.1016/j.jbiosc.2011.11.008.
[28] KOBAYASHI M, HAYASGHI S. Modeling combined effects of temperature and pH on the growth of Zygosa ccharomy ces roux ii in soy sauce mash[J]. Journal of Fermentation and Bioengineering, 1998, 85(6): 638-641. DOI:10.1016/S0922-338X(98)80020-2.
[29] LERTSIRI S, MAUNGMA R, ASSAVANIG A, et al. Roles of the Maillard reaction in browning during Moromi process of Thai soy sauce[J]. Journal of Food Processing and Preservation, 2007, 25(2): 149-162. DOI:10.1111/j.1745-4549.2001.tb00450.x.
Quality and Antioxidant Activity of Raw Sauce Fermented from Rice Dregs with Mixed Starter Cultures
ZHONG Wenxiu, YUAN Jianglan*, LI Chuanwen, KANG Xu, HE Shouchun
(School of Food and Biological and Engineering, Hubei University of Technology, Wuhan 430068, China)
Abstract:In this study, we evaluate the quality and antioxidant activity of raw sauce produced by high-salt liquid-state fermentation of rice dregs with mixed starter cultures. The results showed that all four rice sauces fermented by different mixed starters reached the quality requirements of the Chinese national standard GB 18186-2000 for fi rst-grade soy sauce. The sauce fermented by soy sauce koji containing Aspergillus oryzae and Aspergillus niger plus Zygosaccharomyces rouxii(S4) contained 41.81 g/L total free amino acids. The rice sauces had higher contents of total amino acids, essential amino acids, flavor amino acids and antioxidant amino acids than soy sauce fermented by Aspergillus oryzae (S5). The antioxidant activity of the rice sauces fermented by soy sauce koji containing Aspergillu s o ryzae alone (S3) and both Asperg illusspecies (S4) plus Z yg osaccharomyces rouxii was higher than that of their counterparts (S1 and S2) obtained without theuse of Zygosaccharomyces rouxii. S3 was found to have the strongest antioxidant activity, showing a 175.42- and 2.16-fold increase in hydroxyl and DPPH radical scavenging capacity compared with VC, respectively. The antioxidant activity of rice sauce was strongly infl uenced by fermentation temperature, reaching a peak at 35 and 50 ℃. In conclusion, the quality and antioxidant activity of rice sauce fermented with mixed cultures could be considerably improved compared with single culture fermentation, which is greatly attributed to Zygosaccharomyces rouxii.
Key words:raw sauce produced from rice dregs; quality; antioxidant activity; mixed strains; Zygosaccharomyces rouxii
DOI:10.7506/spkx1002-6630-201706009
中图分类号:TS264.2
文献标志码:A
文章编号:1002-6630(2017)06-0055-07
引文格式:
钟文秀, 袁江兰, 李传雯, 等. 菌种耦合发酵米渣生酱油的品质和抗氧化活性[J]. 食品科学, 2017, 38(6): 55-61.
DOI:10.7506/spkx1002-6630-201706009. http://www.spkx.net.cn
ZHONG Wenxiu, YUAN Jianglan, LI Chuanwen, et al. Quality and antioxidant activity of raw sauce fermented from rice dregs with mixed starter cultures[J]. Food Science, 2017, 38(6): 55-61. (in Chinese with English abstract) DOI:10.7506/ spkx1002-6630-201706009. http://www.spkx.net.cn
收稿日期:2016-05-23
基金项目:国家自然科学基金面上项目(31371741)
作者简介:钟文秀(1990—),女,硕士研究生,研究方向为食品蛋白质发酵。E-mail:18986273776@163.com
*通信作者:袁江兰(1970—),女,副教授,博士,研究方向为食品蛋白质。E-mail:jlyuan1229@163.com