 was reported.
was reported.
DENG Guogang, LI Canpeng *
(School of Chemical Science and Technology, Yunnan University, Kunming 650091, China)
Abstract:In this study, the effects of phosphorylated ovalbumin (PP-OVA) on heat-induced aggregation of native ovalbumin (N-OVA) were investigated. Results showed that PP-OVA effectively suppressed N-OVA aggregation, and the inhibited ability is little weakened when the ionic strength ranged from 10 to 100 mmol/L. Dynamic light scattering analysis showed that the size of the formed N-OVA aggregates decreased in the presence of PP-OVA. Transmission electron micrographs further showed that the N-OVA/PP-OVA mixture exhibited a more uniform size distribution than N-OVA alone. After heat treatment solution of N-OVA, the ζ-potential increased with PP-OVA increased, suggesting that the system became more stable. The result of native-PAGE suggested that the N-OVA/PP-OVA aggregates possess a well-solubility and homogeneity. These results indicate that PP-OVA exhibits a chaperone-like activity under heating, and provide an insight into the possible mechanism through which PP-OVA stabilizes proteins to resist heat-induced N-OVA aggregation.
Key words:molecular chaperone; ovalbumin; phosphorylated ovalbumin; aggregation
DENG Guogang, LI Canpeng. Heat-induced aggregation inhibition properties of phosphorylated ovalbumin through its molecular chaperone-like properties[J]. 食品科学, 2017, 38(5): 66-73. DOI:10.7506/spkx1002-6630-201705011. http://www.spkx.net.cn
DENG Guogang, LI Canpeng. Heat-induced aggregation inhibition properties of phosphorylated ovalbumin through its molecular chaperone-like properties[J]. Food Science, 2017, 38(5): 66-73. (in Chinese with English abstract) DOI:10.7506/ spkx1002-6630-201705011. http://www.spkx.net.cn
Ovalbumin (OVA), a major globular protein component of egg white, exhibits several functional properties, such as foaming and gel forming ability upon heating [1-3]. Native OVA (N-OVA) is a monomeric phosphoglycoprotein that consistsof 385 amino acid residues and has a molecular weight of 45 000 D, an isoelectric point of 4.5, and a denaturation temperature of 84 ℃ at pH 7.0 [4]. Heat-induced denaturation destroys the compact structure of proteins, thereby exposing hydrophobic residues to the surface and exchanging disulfide bonds; this phenomenon results in the formation of intermolecular hydrophobic interactions and disulfide bonds. After denaturation and aggregation, heat-induced N-OVA gels are formed, and this process requires high protein concentration [5]. Meanwhile, aggregation depends on the balance between attractive hydrophobic and electrostatic repulsive interactions [6]. The morphology of aggregates is also affected by several factors, such as pH, heating temperature, ionic strength, and electrostatic repulsive forces [1,7-8]. For example, heating N-OVA aqueous solution produces a transparent solution or turbid suspension because of random aggregation.
A research showed that heat could induce protein denaturation and gelation, but negatively charged N-OVA particles prevented the proteins from coagulating [9]. Several studies investigated the effects of increased protein charges on thermal-induced protein denaturation. Heat-induced N-OVA aggregates transform the solution from turbid to transparent with increasing net charges of N-OVA [10]. Another study suggested that increasing net charges can change the transparency and elasticity of N-OVA gels [3]. Our previous studies further demonstrated that increasing the net charge of N-OVA or egg white protein through phosphorylation by dry heating in the presence of phosphates can improve the heat stability of the solution; this phenomenon could be due to enhanced electrostatic repulsive interactions [11-14]. Consequently, orderly aggregation yields polymers with good solubility, thereby forming a transparent solution.
Many proteins, such as casein, small heat shock proteins, and heme, possess the characteristics of molecular chaperones and have gained increased research attention in food, biology, medical, and pharmaceutical fields [15-20]. Molecular chaperones are a family of proteins that can inhibit protein aggregation under unfavorable conditions (e.g., heat, pressure, oxidatio and reduction) by blocking hydrophobic surfaces exposed from denaturing substrate proteins [21]. For example, casein and small heat shock protein inhibit aggregation through hydrophobic interaction between denatured proteins and molecular chaperones [22-23]. Molecular chaperones should possess the ability to protect from aggregation during protein unfolding under unfavorable conditions, or suppress aggregation during protein refolding from a denatured state, or recover lost biological activity [15,23-25].
Caseins, which belong to the family of phosphoproteins, possess the features of molecular chaperones. Phosphorylated OVA (PP-OVA) obtained using our previously reported method contains many phosphate groups and exhibits special properties, such as easy dissolution and good thermal stability [11]. Caseins, which are neither strictly globular nor fibrous protein [26], do not possess a well-defined secondary or tertiary structure [27]. However, PP-OVA is a globular protein that exhibits a neat secondary structure. Moreover, phosphorylation can enhance the SH-SS exchange reaction and surface hydrophobicity. Considering that caseins contain anionic phosphor-residue clusters and can be a molecular chaperone, we hypothesize whether PP-OVA also exhibits the role of a chaperone to suppress protein aggregation and precipitation under unfavorable conditions. In this paper, we report that modified protein (PPOVA) exhibited chaperone-like activity to inhibit substrate protein (N-OVA) precipitation during thermal treatment. Because molecular chaperone proteins are involved in the folding, transport, assembly, degradation, and stabilization of proteins, PP-OVA is thus expected to be used as a potential molecular chaperone protein in the functions. It is worthy to note that the molecular chaperone-like property of PP-OVA is little affected on the presence of different ionic strength (NaCl concentrations ranging from 10 to 100 mmol/L). Therefore, it is also may be utilized as a molecular chaperone to control protein aggregation in foods.
1.1 Materials and Reagents
N-OVA was purified according to the method in our previous paperas follows [11]: fresh egg white was separated from infertile eggs and homogenized with equivalent deionized water. Then the mixture was precipitated with 2.0 mol/L acetic acid twice at pH 5.5 and 4.5 respectively. The solution was dialyzed for 2 d against 50 mmol/L sodium acetate buffer at pH 4.5. Dialyzed solution was used for column chromatography with CM-cellulose (Whatman International Ltd., Maidstone, Kent, U.K.) equilibrated with 50 mmol/L sodium acetate buffer at pH 4.5, and the OVA fraction was eluted with a 50 mmol/L sodium acetate buffer at pH 4.9. All other reagents were of analytical grade.
1.2 Instruments and Equipment
UV2600A spectrophotometer Unico Ltd. (Shanghai,China); JEM-2100 transmission electron microscopy (TEM) JEOL Ltd. (Tokyo, Japan); Nano-ZS particle size analyzer, Nano-ZS doppler electrophoresis measurements Malvern Instruments (Worcestershire, UK).
1.3 Methods
1.3.1 Preparation of PP-OVA and dry-heated OVA (DH-OVA)
PP-OVA and DH-OVA were prepared according to our previous paper [11]. In brief, N-OVA was dissolved at a concentration of 1 g/100 mL in 0.1 mol/L sodium pyrophosphate buffer at pH 3.0, then the solution was lyophilized. Lyophilized samples were incubated at 85 ℃for 3 d. Dry-heated samples were dissolved and the pH of the solution was adjusted to 7.0 with 1.0 mol/L NaOH. After that the solution was dialyzed and lyophilized to obtain PPOVA. By contrast, DH-OVA was prepared as follows: N-OVA samples were dissolved at 1 g/100 mL in deionized water and the pH of the solution was adjusted to 3.0 with 1.0 mol/L HCl, and then lyophilized. The following operations were under the same conditions as those of PP-OVA.
1.3.2 Determination of phosphorus content
Protein samples were digested in perchloric acid. Phosphorus (P) in the digest was regarded as the total phosphorus of proteins. For the determination of inorganic phosphorus (Pi), 5 mL of 10% trichloroacetic acid was added to the same volume of 1 g/100 mL protein solution, and the solution was centrifuged at 3 000 r/min for 2 h. The phosphorus in the supernatant was regarded as inorganic phosphorus. The phosphorus content was determined according to the method reported previously [28]. The amount of phosphorus bound to proteins was estimated by the difference between the total phosphorus and inorganic phosphorus content.
1.3.3 Measurement of sulfhydryl thiol (SH) groups
Free SH groups were determined using 5,5’-dithiobis-(2-nitrobenzoic acid) (DTNB) according to the procedure of Ellman [29]. Briefly, to 1 mL of 0.1 g/100 mL protein solution were added 1 mL of 0.1 mol/L Tris-glycine buffer (pH 8.0) containing 0.01 mol/L ethylene diamine tetraacetic acid (EDTA) (for surface SH groups) and the same buffer containing 5% dodecyl sulfate, sodium (SDS) (for total SH groups). After incubation at 40 ℃ for 30 min, 50 μL of DTNB solution (4 mg in 1 mL of 0.1 mol/L Tris-glycine buffer, pH 8.0) was added and then incubated at 25 ℃ for 10 min. Absorbance was read at 412 nm. A molar extinction coefficient of 13 600 L/(mol·cm) at 412 nm of 2-nitro-5-mercaptobenzoic acid was used for calculation of the modified sulfhydryl groups in the OVA molecule using DTNB. The SH group content was calculated as moles of SH per mole of OVA.
1.3.4 Determination of pH and temperature effect on N-OVA aggregation
N-OVA solutions (0.2 g/100 mL protein) were prepared by dissolveing in 50 mmol/L sodium phosphate buffer at various pH values in sealed glass vials. The N-OVA solutions were heated in a water bath at 65–90 ℃ for 20 min. Vials were removed and cooled immediately in a ice water for 5 min. Protein aggregation extent was determined by solution turbidity as optical density (OD) at 400 nm using UV spectrophotometer. Based on these results, a determinate pH of 6.0 of sodium phosphate buffer for all subsequent experiments [20].
1.3.5 Concentration-dependent effect of PP-OVA on N-OVA aggregation
PP-OVA or DH-OVA (0–0.2 g/100 mL) and N-OVA (0.2 g/100 mL) were dissolved in 50 mmol/L sodium phosphate buffer at pH 6.0 in sealed glass vials. The mixed solutions were heated in a water bath at 90 ℃ for 20 min before they were cooled immediately in an ice water bath. Then OD was determined at 400 nm.
1.3.6 Native polyacrylamide gel electrophoresis (native-PAGE)
Native-PAGE was consulted a common method [30]. Briefly, native-PAGE without SDS and with or without reducing agent was performed in 4 g/100 mL stacking gel and 7.5 g/100 mL separating gel. N-OVA (0.2 g/100 mL), PP-OVA (0.2 g/100 mL) and N-OVA (0.2 g/100 mL) added or not added PP-OVA (0.2 g/100 mL) were dissolved respectively in 50 mmol/L sodium phosphate buffer at pH 6.0 in sealed glass vials. Unheating or heating was consistent with previous processing method. The samples were centrifuged at 5 000 r/min for 5 min after heating to obtain supernate. The 15 μL sample solutions mixed equivalent loading buffer (0.5 mol/L, pH 6.8 Tris-HCl, 0.2 g/100 mL bromophenol blue, 30% (V/V) glycerol ) were loaded on gel and separated at 100 V. After that, the gel was stained and destained overnight.
1.3.7 Transmission electron microscopy (TEM)
The microstructure of heat-induced N-OVA, PP-OVA and N-OVA/PP-OVA mixture were acquired with TEM. Using an acceleration voltage of 200 kV. All samples were obtained by 0.2 g/100 mL proteins were dissolved respectively in 50 mmol/L phosphate buffered saline (PBS) at pH 6.0, heating at 90 ℃ for 20 min, ice bath and diluting to 5-fold.1.3.8 Particle size
Particle size analysis was carried out at 25 ℃ using a Nano-ZS particle size analyzer. The sample solutions were diluted to 1:50 to avoid multiple scattering phenomena due to interparticle interaction [31]. Then the diluted sample was transferred into the polystyrene curet for size determination, the cumulative method was used to fi nd the average diameter. The particle size distribution was reported as polydispersity index (PDI) from the DLS analyzer, in which PDI <1 represented the good polydispersion.
1.3.9 ζ-Potential
ζ-Potential was determined from laser doppler electrophoresis measurementsat 25 ℃ using the Smoluchowski model. The sample solutions were diluted to 1:50 and were analyzed in folded capillary cells.
1.3.10 The effect of NaCl on the inhibiting ability
The mixed solutions of N-OVA (0.2 g/100 mL) and PP-OVA or DH-OVA (0.2 g/100 mL) in the buffer (pH 6.0) containing various concentrations of NaCl (0–100 mmol/L) were heated at 90 ℃ for 20 min and placed in an ice bath for 5 min. Protein aggregation extent was determined by solution turbidity as optical density (OD) at 400 nm using UV spectrophotometer.
1.4 Date analysis
All experiments were performed in triplicate, and
was reported.
2.1 Characteristics of PP-OVA

Fig. 1 Native-PAGE of N-, DH- and PP-OVA
Table 1 Some characteristics of N-, DH- and PP-OVA

Note: *. The difference was signif i cant (P<0.05); **. The difference was very signif i cant (P<0.01). The same below.
ProteinP content/%SH group content/(mol/mol) SurfaceTotal N-OVA0.121.20 ± 0.013.56 ± 0.01 DH-OVA0.121.51 ± 0.02 **3.46 ± 0.02 PP-OVA 1.91 **2.31 ± 0.01 **2.97 ± 0.04 **
N-OVA was phosphorylated by dry heating at pH 3.0 and 85 ℃ for 3 d. The P content of the phosphorylated N-OVA is shown in Table 1. Generally, the P content of N-OVA, which is naturally phosphorylated at serine -68 and/or -344, is 0.12% [32]. In this study, the P content of phosphorylated N-OVA was 1.91%, which is higher than that reported in previous studies [11,13]. The discrepancy in the results could be due to differences in the experimental conditions employed. As such, we determined the inf l uence of pH on phosphorylation by using pH 3.0, which has been reported to present an optimal effect. The native-PAGE patterns of N-, DH- and PP-OVA are shown in Fig. 1. The mobility of N-OVA and DH-OVA did not change, whereas that of PP-OVA increased. This fi nding indicates that a high level of negatively charged phosphate groups on PP-OVA can improve protein mobility, which is consistent with that reported in previous research [11].
Changes in the surface and total SH group contents in OVA are shown in Table 1. The surface SH content of DH-OVA and PP-OVA increased, whereas the total SH group content of DH-OVA and PP-OVA decreased. Hence, phosphorylation enhanced the decrease in total SH group content. These results indicate the exposure of SH groups to surface OVA molecules, and subsequent SH-SS exchange reactions and SH oxidation may simultaneously occur by dry heating [11-12].
2.2 Effect of pH and temperature on N-OVA aggregation

Fig. 2 Temperature-dependent changes of turbidity of N-OVA at different pH values
The turbidity of 0.2 g/100 mL N-OVA solution (pH 5.5–8.0) after heating at 65 to 90 ℃ is shown in Fig. 2. Theturbidity of the heated solutions did not differ at pH≥6.5, whereas large aggregates started to appear at pH 5.5. The N-OVA solution remained homogeneous at pH 6.0. Thus, pH 6.0 was used as the suitable condition for further study. The minimum turbidity was observed at pH 6.0 when solutions were heated at 70 ℃. Therefore, temperature within the range of 75–90 ℃ was used to create a stressed heating condition for subsequent experiments [20].
2.3 Effect of PP-OVA on N-OVA aggregation
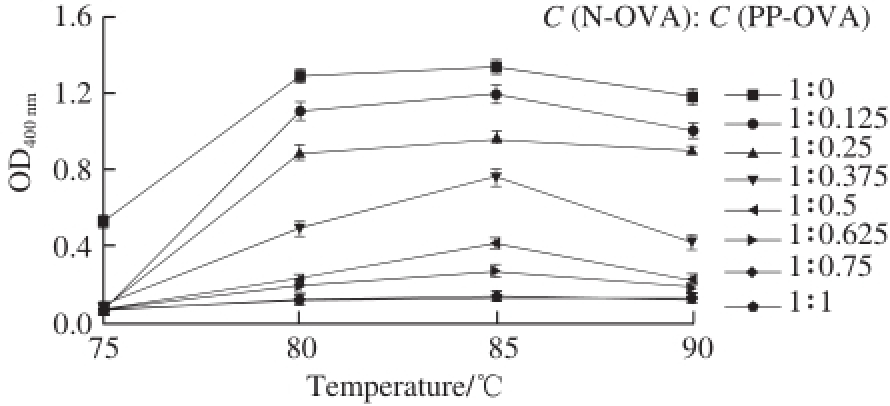
Fig. 3 Temperature-dependent changes in the turbidity of N-OVA with increasing PP-OVA concentration at pH 6.0
The effect of 0.025–0.200 g/100 mL PP-OVA on N-OVA aggregation was determined by measuring turbidity at 400 nm [16]. The maximum value for turbidity (OD) was 1.29 ± 0.04 in the absence of PP-OVA (Fig. 3). The addition of 0.2 g/100 mL PP-OVA reduced turbidity to only 0.13 ± 0.005 after heating at 85 ℃. Heat-induced aggregation of N-OVA was also suppressed at a C (N-OVA): C (PP-OVA) of 1: 0.75. Approximately 90% of OVA (0.1 g/100 mL) aggregation was suppressed, whereas whole casein completely inhibited OVA aggregation at 1:1 (m/m) concentration and in the presence of β-casein at about 0.2 g/100 mL [16]. The turbidity of N-OVA decreased with increasing PP-OVA concentration. Thus, PPOVA can remarkably inhibit N-OVA aggregation.
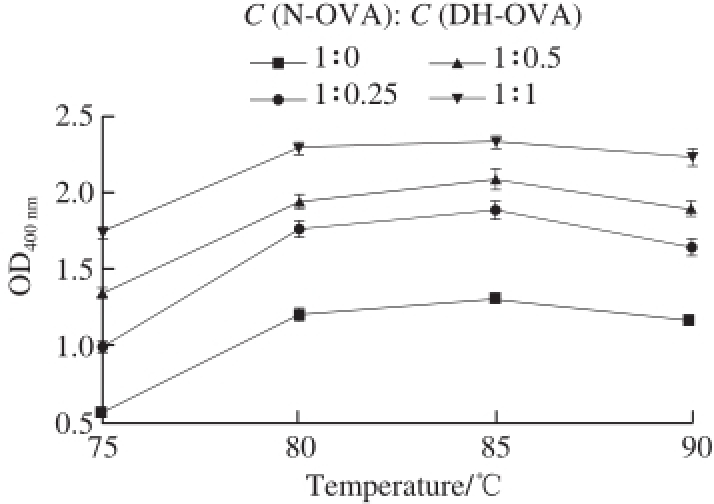
Fig. 4 Temperature-dependent changes in the turbidity of N-OVA with increasing DH-OVA concentration at pH 6.0
Interestingly, the maximum turbidity was achieved at 85 ℃, regardless of pH (Fig. 2) or PP-OVA concentration (Fig. 3). This phenomenon could be related to differences in surface hydrophobicity caused by temperature changes [20]. In this study, the effect of DH-OVA on N-OVA aggregation was investigated. The turbidity of N-OVA did not decrease but increase with increasing DH-OVA concentration (Fig. 4). This phenomenon suggests that DH-OVA could not inhibit N-OVA aggregation during thermal denaturation. Fig. 3 shows the potential of PP-OVA for altering heat-induced aggregation of N-OVA solutions. The effects of PP-OVA were concentration dependent, such that PP-OVA (0.025–0.200 g/100 mL) incrementally decreased the turbidity of N-OVA (0.2 g/100 mL) at pH 6.0 when heated at 90 ℃. Ordered secondary structures were observed for spherical N-OVA, which can easily precipitate under heating. However, PP-OVA itself not only did not precipitate but also inhibit the heat-induced precipitation of N-OVA.
2.4 Changes in physicochemical characteristics of N-OVA/ PP-OVA mixture
Table 2 Changes in the particle size and ζ-potential of N-OVA with increasing PP-OVA concentration
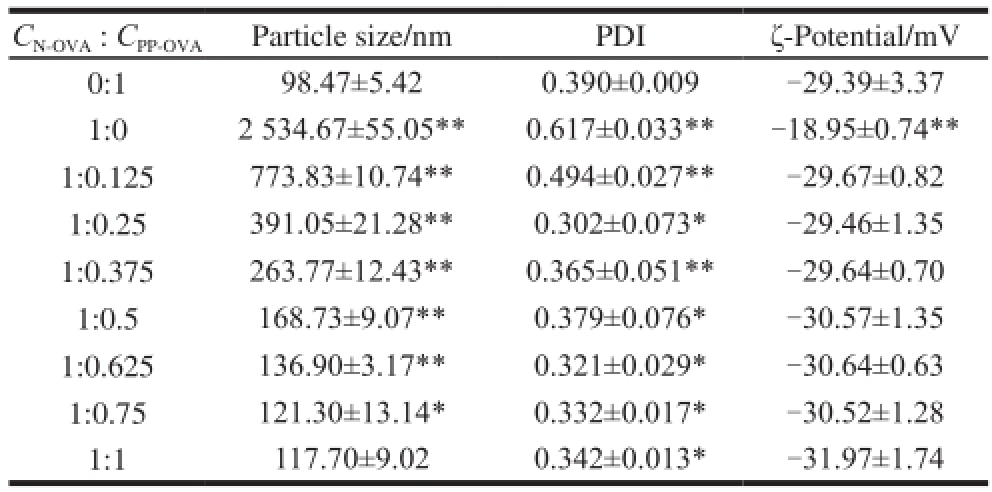
Note: All sample solutions were obtained by heating at 90℃, pH 6.0, ice bath and diluting to 50-fold.
C N-OVA: C PP-OVAParticle size/nmPDIζ-Potential/mV 0:198.47±5.420.390±0.009-29.39±3.37 1:02 534.67±55.05**0.617±0.033**-18.95±0.74** 1:0.125773.83±10.74**0.494±0.027**-29.67±0.82 1:0.25391.05±21.28**0.302±0.073*-29.46±1.35 1:0.375263.77±12.43**0.365±0.051**-29.64±0.70 1:0.5168.73±9.07**0.379±0.076*-30.57±1.35 1:0.625136.90±3.17**0.321±0.029*-30.64±0.63 1:0.75121.30±13.14*0.332±0.017*-30.52±1.28 1:1117.70±9.020.342±0.013*-31.97±1.74
The particle size, ζ-potential, and PDI of N-OVA solutions with PP-OVA or without PP-OVA are presented in Table 2. The particle sizes of PP-OVA and N-OVA were (98.47±5.42) and (2 534.67±55.05) nm, respectively. The particle size of N-OVA decreased with increasing PP-OVA concentration and was within the range of 2 534.67 nm to 117.7 nm by adding PP-OVA (0.2 g/100 mL). This characteristic was due to enhanced electronic repulsion caused by increased phosphate groups. In another study, the protein particle size of whey protein isolate/micellar casein (1:1, m/m) significantly increased from an initial diameter of 165.5 nm in the unheated mixture to 272 nm after heating at 85 ℃ (pH 6.0) for 10 min [33]. Hence, the electronic repulsion of the N-OVA/PP-OVA complex was increased, whereas N-OVA aggregation was reduced. This phenomenon resulted in the smaller particle size of the mixture compared with that of N-OVA. Low PDI indicates the presence of ahomogeneous dispersion. The results showed that PDI ranged from 0.302 to 0.617, but all values were less than 1, showing good polydispersion of the N-OVA/PP-OVA complex.
ζ-Potential, i.e., surface charge, can influence the stability of nanoparticles through electrostatic repulsion. The ζ-potentials of PP-OVA and N-OVA were −29.39 and −18.95 mV, respectively. The ζ-potential of the N-OVA/PP-OVA complex ranged from −18.95 to −31.97 mV. Hence, N-OVA had lower ζ-potential than the N-OVA/PP-OVA mixture. After heating of the mixed N-OVA and PP-OVA, N-OVA aggregation and precipitation were suppressed because of the abundant negative charges of PP-OVA. ζ-Potential was not affected by pH after heating [34]. The increase in ζ-potential induced by adding PP-OVA was higher for N-OVA because nanoparticle aggregation decreased with increasing ζ-potential caused by electric repulsion. This behavior indicates the high physical stability of the N-OVA/PP-OVA complex. The aggregates [10]or transparent gels [3]were also significantly affected by the net charge of OVA within a wide range of charges derived from methylation or succinylation.
2.5 TEM of N-OVA, PP-OVA and N-OVA/PP-OVA mixture

Fig. 5 TEM of heated OVA samples
The typical morphologies of N-OVA, PP-OVA and N-OVA/PP-OVA mixture after heating are shown in Fig. 5. After heating, large and amorphous N-OVA, with massive texture and a size of > 2 μm, precipitated (Fig. 5a). PP-OVA had a round shape and a particle size of approximately 107–136 nm (Fig. 5b). These results could be due to the inter-repulsion of small-sized PP-OVA with high surface charges. The N-OVA/PP-OVA mixture (1:1, m/m) contained mainly smaller, elongated, homogenous aggregates, with a size of 115–137 nm (Fig. 5c). The TEM morphological results are consistent with those of particle size distribution shown in Table 2. This phenomenon indicates that PP-OVA effectively inhibited N-OVA precipitation under heating, which could be attributed to the different structures and inf l uence of electrostatic interaction. Thus, PP-OVA can be potentially applied as a molecule chaperon.
2.6 Electrophoresis
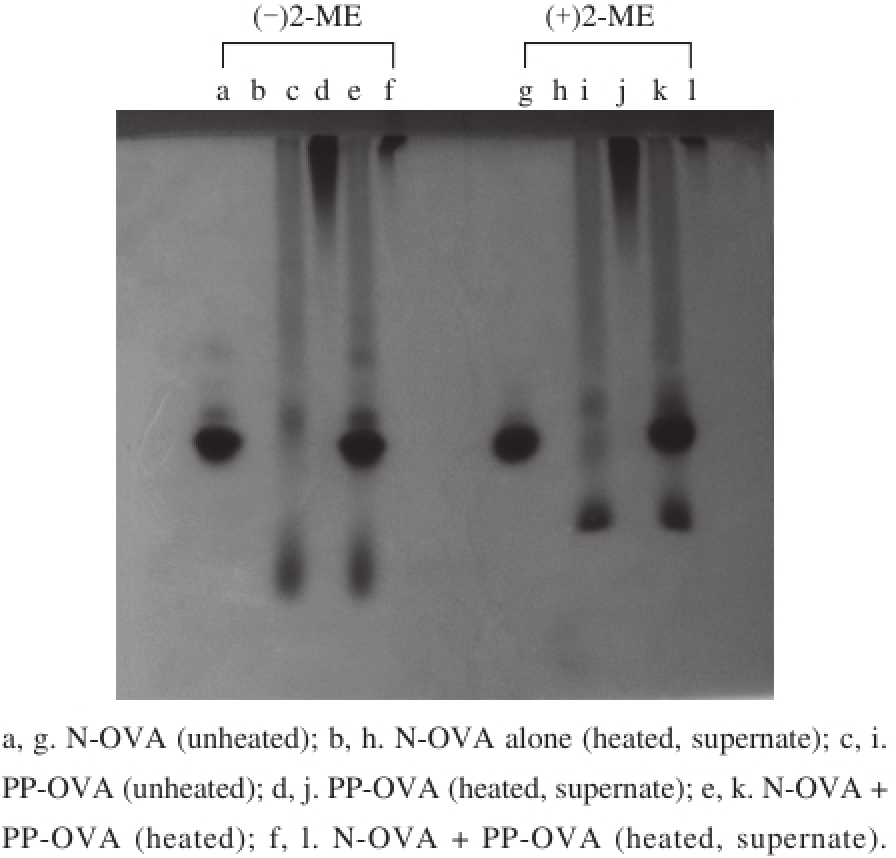
Fig. 6 Native-PAGE patterns with (+) and without (-) 5% of 2-mercaptoethanol (2-ME)
High levels of negatively charged phosphate groups on OVA cause increased mobility [11], which can be distinguished from Fig. 1. A possible mechanism for the suppression of PPOVA against N-OVA aggregation was determined by heating N-OVA, PP-OVA, and N-OV/PP-OVA mixture at 90 ℃ for 20 min. After which, the solutions were placed in an ice bath. The supernatant fraction was obtained by centrifugation and applied to native-PAGE. Unheated samples were also applied to native-PAGE for comparison. The results are shown in Fig. 6. The bands (Fig. 6a and c) of N-OVA and PPOVA are consistent with those presented in Fig. 1. Unheated N-OVA/PP-OVA mixture retained its original characteristics (Fig. 6e and k). The dimer and oligomer of N-OVA emerged in the absence of 2-mercaptoethanol (2-ME), but the bands of dimer and oligomer disappeared in the presence of 2-ME (Fig. 6a, g, e, k). When N-OVA solution was heated, all samples containing N-OVA produced insoluble aggregates, and the supernatant fraction did not exhibit any bands at the position corresponding to unheated N-OVA (Fig. 6b and h). When PP-OVA solution was heated, PP-OVA produced soluble and homogeneous aggregates, and bands (Fig. 6d and j) were observed at the top of the separating gel. The bands (Fig. 6f and l) of soluble aggregates were further detected at the top of separating gel in the N-OVA solutionin the presence of PP-OVA. However, the bands of N-OVA and PP-OVA disappeared completely. These results suggest that PP-OVA may have interacted with N-OVA because of electrostatic interaction during heating. Thus, PP-OVA probably created a nonreactive barrier by placing itself between the heat-denatured N-OVA molecules. So PP-OVA may have suppressed N-OVA interaction, resulting in soluble aggregates.
2.7 Effect of NaCl on the suppressive ability of PP-OVA or DH-OVA

Fig. 7 Inf l uence of NaCl concentration on OD at 400 nm (A) and appearance (B)
The effect of NaCl on the ability of PP-OVA or DH-OVA to suppress heat-induced aggregation of N-OVA was investigated. The results of turbidity development and appearance are shown in Fig. 7A and B, respectively. The turbidity (400 nm) of N-OVA and DH-OVA solutions slowly increased with increasing NaCl concentration, whereas that of PP-OVA solution was low and remained transparent at all NaCl concentrations. Moreover, the turbidity of the mixture of N-OVA and DH-OVA at 400 nm was enhanced with increasing NaCl concentration. This result indicates that DH-OVA does not affect N-OVA aggregation. Furthermore, the turbidity of the mixed N-OVA and PP-OVA solution slightly increased with increasing NaCl concentration. The maximum value for turbidity was 0.81 at 400 nm and 100 mmol/L NaCl. Thus, the ability of PP-OVA to suppress N-OVA aggregation was not affected by the presence of NaCl, which maybe due to the repulsion caused by introduced phosphate groups in PP-OVA. Increasing the salt concentration of the protein solution affected the aggregate morphology by shielding the exposed charges of the protein [10]. Other experiments showed that casein suppression of protein aggregation weakened with increasing NaCl concentration, which was due to the severely reduced casein charge [25,33]. Interestingly, the suppression ability of N-OVA aggregation by PP-OVA was hardly affected in the presence of NaCl. The result suggested that the interaction other than repulsion interaction caused by phosphate group, PP-OVA may also play the role of suppressing aggregation of N-OVA. We previously reported that PP-OVA exhibits a molten globular protein structure, which binds to N-OVA molecules and therefore form stable N-OVA/PP-OVA aggregates. Furthermore, we have demonstrated that the surface hydrophobicity of PP-OVA is much larger than that of DH-OVA [11], which may affect the aggregation of OVA. Therefore, the increased surface hydrophobicity in PP-OVA is also considered to be one of the reasons why PP-OVA is effective in anti-aggregation.
This work shows that PP-OVA could inhibit N-OVA aggregation during thermal treatment, and this behavior was attributed to a chaperone-like activity. The minimum concentration ratio for N-OVA to PP-OVA was 1:0.75 (m/m). N-OVA solution was transparent and homogeneous in the present of PP-OVA after heating. Several characteristics, such as decreased particle size and increased ζ-potential, were altered, resulting in the good dispersion and stability of the N-OVA/PP-OVA mixture. Furthermore, the inf l uence of ionic strength on the inhibitory effect of PP-OVA against N-OVA precipitation was weak. The results show that PPOVA can function as a chaperone that could suppress matrix protein aggregation under heating. It may be anticipated that PP-OVA may recover of lost biological activity and be applied in food and other fi elds.
References:
[1] DOI E, KITABATAKE N. Structure of glycinin and ovalbumin gels[J]. Food Hydrocolloids, 1989, 3(4): 327-337. DOI:10.1016/ S0268-005X(89)80043-8.
[2] HAGOLLE N, RELKIN P, DALGLEISH D G, et al. Transition temperatures of heat-induced structural changesin ovalbumin solutions at acid and neutral pH[J]. Food Hydrocolloids, 1997, 11(3): 311-317. DOI:10.1016/S0268-005X(97)80061-6.
[3] MINE Y. Laser light scattering study on the heat-induced ovalbumin aggregates related to its gelling property[J]. Journal of Agricultural and Food Chemistry, 1996, 44(8): 2086-2090. DOI:10.1021/JF960153Z.
[4] MINE Y. Recent advances in the understanding of egg white protein functionality[J]. Trends in Food Science & Technology, 1995, 6(7): 225-232. DOI:10.1016/S0924-2244(00)89083-4.
[5] ARNTFIELD S D, MURRAY E D, ISMOND M A H. Role of the thermal denaturation-aggregation relationship in determining the rheological properties of heat induced networks for ovalbumin and vicilin[J]. Journal of Food Science, 1989, 54(6): 1624-1631. DOI:10.1111/j.1365-2621.1989.tb05175.x.
[6] ALTING A C, WEIJERS M, HOOG E H A D, et al. Acid-induced cold gelation of globular proteins: effects of protein aggregate characteristics and disulfide bonding on rheological properties[J]. Journal of Agricultural and Food Chemistry, 2004, 52(3): 623-631. DOI:10.1021/jf034753r.
[7] KITABATAKE, N, KINEKAWA, Y I. Turbidity measurement of heated egg proteins using a microplate system[J]. Food Chemistry, 1995, 54(2): 201-203. DOI:10.1016/0308-8146(95)00032-E.
[8] ARNTFIELD S D, BERNATSKY A. Characteristics of heat-induced networks for mixtures of ovalbumin and lysozyme[J]. Journal of Agricultural and Food Chemistry, 1993, 41(12): 2291-2295. DOI:10.1021/jf00036a013.
[9] HU J H, YU S Y, YAO P. Stable amphoteric nanogels made of ovalbumin and ovotransferrin via self-assembly[J]. Langmuir, 2007, 23(11): 6358-6364. DOI:10.1021/la063419x.
[10] WEIJERS M, BROERSEN K, BARNEVELD P A, et al. Net charge affects morphology and visual properties of ovalbumin aggregates[J]. Biomacromolecules, 2008, 9(11): 3165-3172. DOI:10.1021/ bm800751e.
[11] LI C P, HAYASHI Y, SHINOHARA H, et al. Phosphorylation of ovalbumin by dry-heating in the presence of pyrophosphate: effect on protein structure and some properties[J]. Journal of Agricultural and Food Chemistry, 2005, 53(12): 4962-4967. DOI:10.1021/jf047793j.
[12] YIN C Y, YANG L, ZHAO H, et al. Improvement of antioxidant activity of egg white protein by phosphorylation and conjugation of epigallocatechin gallate[J]. Food Research International, 2014, 64: 855-863. DOI:10.1016/j.foodres.2014.08.020.
[13] LI C P, SALVADOR A S, IBRAHIM H R, et al. Phosphorylation of egg white proteins by dry-heating in the presence of phosphate[J]. Journal of Agricultural and Food Chemistry, 2003, 51(23): 6808-6815. DOI:10.1021/jf030043+.
[14] LI C P, HAYASHI Y, ENOMOTO H, et al. Phosphorylation of proteins by dry-heating in the presence of pyrophosphate and some characteristics of introduced phosphate groups[J]. Food Chemistry, 2009, 114(3): 1036-1041. DOI:10.1016/j.foodchem.2008.10.066.
[15] HE J S, ZHU S M, MU T H, et al. αs-casein inhibits the pressureinduced aggregation of β-lactoglobulin through its molecular chaperonelike properties[J]. Food Hydrocolloids, 2011, 25(6): 1581-1586. DOI:10.1016/j.foodhyd.2011.01.008.
[16] KHODARAHMI R, BEYRAMI M, SOORI H. Appraisal of casein’s inhibitory effects on aggregation accompanying carbonic anhydrase refolding and heat-induced ovalbumin fi brillogenesis[J]. Archives of Biochemistry and Biophysics, 2008, 477(1): 67-76. DOI:10.1016/ j.abb.2008.04.028.
[17] KHODARAHMI R, SOORI H, KARIMI S A. Chaperone-like activity of heme group against amyloid-like fibril formation by hen egg ovalbumin: possible mechanism of action[J]. International Journal of Biological Macromolecules, 2009, 44(1): 98-106. DOI:10.1016/ j.ijbiomac.2008.10.011.
[18] NIFOROU K, CHEIMONIDOU C, TROUGAKOS I P. Molecular chaperones and proteostasis regulation during redox imbalance[J]. Redox Biology, 2014, 2: 323-332. DOI:10.1016/j.redox.2014.01.017.
[19] TREWEEK T M, MORRIS A M, CARVER J A. Intracellular protein unfolding and aggregation: the role of small heat-shock chaperone proteins[J]. Australian Journal of Chemistry, 2003, 56(5): 357-367. DOI:10.1071/CH03031.
[20] YONG Y H, FOEGEDING E A. Effects of caseins on thermal stability of bovine β-lactoglobulin[J]. Journal of Agricultural and Food Chemistry, 2008, 56(21): 10352-10358. DOI:10.1021/jf801658u.
[21] FRYDMAN J. Folding of newly translated proteins in vivo: the role of molecular chaperones[J]. Annual Review of Biochemistry, 2001, 70: 603-647. DOI:10.1146/annurev.biochem.70.1.603.
[22] MACRAE T H. Structure and function of small heat shock/alphacrystallin proteins: established concepts and emerging ideas[J]. Cellular and Molecular Life Sciences, 2000, 57(6): 899-913. DOI:10.1007/PL00000733.
[23] SAKONO M, MOTOMURA K, MARUYAMA T, et al. Alpha casein micelles show not only molecular chaperone-like aggregation inhibition properties but also protein refolding activity from the denatured state[J]. Biochemical and Biophysical Research Communications, 2011, 404(1): 494-497. DOI:10.1016/j.bbrc.2010.12.009.
[24] MANNA T, SARKAR T, PODDAR A, et al. Chaperone-like activity of tubulin. binding and reactivation of unfolded substrate enzymes[J]. The Journal of Biological Chemistry, 2001, 276(43): 39742-39747. DOI:10.1074/jbc.M104061200.
[25] MATSUDOMI N, KANDA Y, YOSHIKA Y, et al. Ability of αscasein to suppress the heat aggregation of ovotransferrin[J]. Journal of Agricultural and Food Chemistry, 2004, 52(15): 4882-4886. DOI:10.1021/jf030802o.
[26] ALAIMO M H, FARRELL H M J, GERMANN M W. Conformational analysis of the hydrophobic peptide αs1-casein (136-196)[J]. Biochimica et Biophysica Acta, 1999, 1431(2): 410-420. DOI:10.1016/ S0167-4838(99)00061-8.
[27] BHATTACHARYYA J, DAS K P. Molecular chaperone-like properties of an unfolded protein, αs-casein[J]. Journal of Biological Chemistry, 1999, 274(22): 15505-15509. DOI:10.1074/jbc.274.22.15505.
[28] CHEN P S, TORIBARA T Y, WARNER H. Microdetermination of phosphorus[J]. Analytical Chemistry, 1956, 28: 1756-1758. DOI:10.1021/ac60119a033.
[29] ELLMAN G T. Tissue sulphydryl groups[J]. Archives of Biochemistry and Biophysics, 1959, 82(1): 70-75. DOI:10.1016/0003-9861(59)90090-6.
[30] LAEMMLI U K. Cleavage of structure proteins during the assembly of the head of bacteriophage T 4[J]. Nature, 1970, 227: 680-685. DOI:10.1038/227680a0.
[31] TAN C, XU J, ERIC K, et al. Dual effects of chitosan decoration on the liposomal membrane physicochemical properties as affected by chitosan concentration and molecular conformation[J]. Journal of Agricultural and Food Chemistry, 2013, 61(28): 6901-6910. DOI:10.1021/jf401556u.
[32] NISBET A D, SAUNDRY R H, MORI A J G, et al. The complete amino acid sequence of hen ovalbumin[J]. European Journal of Biochemistry, 1981, 115(2): 335-345. DOI:10.1111/j.1432-1033.1981.tb05243.x.
[33] O’KENNEDY B T, MOUNSEY J S. Control of heat-induced aggregation of whey proteins using casein[J]. Journal of Agricultural and Food Chemistry, 2006, 54(15): 5637-5642. DOI:10.1021/ jf0607866.
[34] KEHOE J J, FOEGEDING E A. The characteristics of heatinduced aggregates formed by mixtures of β-lactoglobulin and β-casein[J]. Food Hydrocolloids, 2014, 39: 264-271. DOI:10.1016/ j.foodhyd.2014.01.019.
磷酸化卵白蛋白通过其分子伴侣样特性抑制蛋白质热致聚合
邓国刚,李灿鹏
*
(云南大学化学科学与工程学院,云南 昆明 6 5 0 0 9 1)
摘 要:实验研究磷酸化卵白蛋白(phosphorylated ovalbumin,PP-OVA)对原始卵白蛋白(native ovalbumin,N-OVA)的热致聚合影响。结果表明,PP-OVA能有效抑制N-OVA的聚合,并且抑制能力在离子强度为10~100 mmol/L的范围内轻微减弱。动态光散射分析显示,PP-OVA的存在使N-OVA聚合物的粒径减小。透射电子显微镜结果进一步显示PP-OVA/N-OVA聚合体比N-OVA单独存在展现出更均匀的颗粒大小分布。热处理N-OVA的ζ-电位随PP-OVA浓度的增加而增大,表明体系变得更稳定。非变性聚丙烯酰氨凝胶电泳表明PP-OVA/N-OVA聚合体具有较好的溶解性和均一性。在加热条件下,PP-OVA显示了分子伴侣样活性。据此本研究并提出了PP-OVA稳定蛋白去抵抗热致聚合的可能机制。
关键词:分子伴侣;卵白蛋白;磷酸化卵白蛋白;聚合
中图分类号:Q518.4
文献标志码:A
文章编号:1002-6630(2017)05-0066-08
引文格式:
收稿日期:2016-07-14
基金项目:国家自然科学基金地区科学基金项目(21565029);云南大学第三批青年英才培育计划项目(WX069051)
作者简介:邓国刚(1990—),男,博士研究生,主要从事卵白蛋白的改性方面研究。E-mail:348919912@qq.com
DOI:10.7506/spkx1002-6630-201705011
*通信作者:李灿鹏(1974—),男,教授,博士,主要从事功能食品、食品安全与检测方面研究。E-mail:lcppp1974@sina.com