表 1 基础饲料及特征饲料的营养成分
Table 1 Nutrient contents of basic diet and characteristic diets

宋 奇1,艾连中2,鲁红岩1,郝 丹1,陈 燏1,孙 荟1,兰海静1,张祖韬3,李全阳1,*
(1.广西大学轻工与食品工程学院,广西 南宁 530004;2.上海理工大学医疗器械与食品学院,上海 200093;3.广西石埠乳业有限责任公司,广西 南宁 530021)
摘 要:为了验证巴马长寿饮食模式的抗衰老作用,通过给小鼠皮下注射D-半乳糖(500 mg/(kg·d))制备衰老模型,根据本团队前期研究结果,选取巴马长寿老人饮食中具有代表性的营养特征(能量限制及VA、大豆异黄酮、膳食纤维、微量元素铁、锰、钴、硒的摄入),分别设计了能限组、胡豆组、纤维组、元素组和复合组饮食,并以自由采食的方式饲喂小鼠,连续8 周。对小鼠血清、肝脏和脑组织中丙二醛(malondialdehyde,MDA)水平、血清和脑组织中总超氧化物歧化酶(total superoxide dismutase,T-SOD)活力、肝脏和脑组织中总抗氧化能力(total antioxidant capacity,T-AOC)进行测定。结果表明:与衰老组小鼠相比,5 种特征饮食都能抑制小鼠体内MDA的蓄积,提高抗氧化酶活力和T-AOC;通过正向化和无量纲化处理,发现复合组饮食抗氧化应激效果最强,能显著抑制衰老小鼠血清、肝脏和脑组织中MDA蓄积(P<0.05),使血清中T-SOD活力增强12.33%(P<0.05),并使肝脏中T-AOC显著提高35.35%(P<0.05)。5 种模式综合量化评价的顺序为:复合组>胡豆组>能限组>纤维组>元素组。说明巴马长寿饮食模式对D-半乳糖致衰老小鼠有明显的抗氧化应激作用,这也预示着其具有较显著的抗衰老作用效果和良好的开发潜力。
关键词:巴马长寿饮食模式;抗氧化应激;D-半乳糖;衰老模型
衰老是极复杂的过程,受遗传因素和环境因素的共同影响。自由基损伤学说是解释衰老的理论之一,其认为自由基和活性氧在机体内的堆积会导致细胞和组织的损伤,造成氧化应激并引起衰老和不正常的细胞死亡[1]。实际上,有研究表明老年性疾病和神经退行性疾病的发生也伴随着氧化应激[2],因此机体氧化应激水平,如丙二醛(malondialdehyde,MDA)含量、抗氧化酶活力等,也成为衡量抗衰老的重要指标[3],而降低机体氧化应激水平、维持氧化还原平衡对人们健康长寿也有着重要意义[4]。在衰老研究方面,因为高剂量的D-半乳糖难以完全代谢造成其在机体内的蓄积,进而诱导体内氧化应激,可以模拟小鼠的自然衰老,啮齿类动物长期注射D-半乳糖也成为抗衰老研究的动物模型[5],同时其也广泛应用于健康食品的检测。
饮食是对衰老影响最大的环境因素之一[6]。世界卫生组织和美国疾病控制预防中心预估,健康的饮食能有效预防冠心病、II型糖尿病、心血管疾病和癌症等老龄化疾病的发生[7-8]。对饮食营养和健康长寿的研究也表明,合理的饮食能有效延缓衰老[9],如地中海饮食能有效降低心血管疾病和癌症的发生风险[10]。此外,饮食干预如能量限制等,也被证明是改善模型动物衰老特征、延长寿命强有力的手段[11-12]。而对长寿地区饮食营养的研究也表明,长寿地区的饮食结构和营养特征对当地人群的健康长寿有着积极作用[13-14]。
广西河池巴马县是世界知名的长寿之乡,长寿现象凸显。根据2010年中国第6次人口普查资料显示[15],巴马县的百岁人口比例达到36.50/10万 人,远高于联合国对长寿乡的定义标准(百岁人口比例7.50/10万 人)[16]。在巴马地区长寿现象的研究中,对遗传因素和地理环境因素的探索报道较多,本课题组着眼于饮食营养、肠道微生态和代谢组学分析,前期研究中已对巴马长寿地区的饮食进行了调查,发现其是以粥类、玉米、大豆等粗杂粮类、深色蔬菜类、家畜肉类食物摄入为主的膳食纤维多糖饮食模式[17-18];对饮食营养特征的调查也表明,高膳食纤维含量、高VA含量和低能量的摄入特征有助于长寿[19];对巴马百岁老人代谢特征、指甲和饮水中的元素特征及其与营养素摄入水平的相关性分析表明,能量限制、VA、膳食纤维和铁、锰、钴、硒等微量元素的较高摄入量是巴马长寿老人饮食中具有代表性的营养特征,可能与延缓衰老密切相关[20-21]。在上述研究的基础上,试图建立几种巴马长寿饮食模式,并利用动物模型加以验证,以期从饮食的角度探索长寿现象的形成。
为此,本研究选取巴马长寿老人饮食中的特征营养素和营养特征,以D-半乳糖致衰老小鼠为研究对象,通过对小鼠生长特性和氧化应激水平的初步评估,以期探究巴马长寿饮食模式在延缓衰老中的作用。
8 周龄SPF级雄性昆明小鼠84 只,体质量(36±2)g,购自广西医科大学实验动物中心(实验动物生产许可证号:SCXK(桂)2014-002),饲养温度控制在(24±2)℃,相对湿度(60±10)%,12 h昼夜交替,自由饮食饮水。
基础饲料购自北京科澳协力饲料有限公司,符合GB 13078—2017《饲料卫生标准》和GB 14924.3—2010《实验动物 配合饲料营养成分》;β-胡萝卜素 南京都莱生物技术有限公司;大豆异黄酮 上海源叶生物科技有限公司;大豆膳食纤维 平顶山金晶生物科技有限公司;玉米膳食纤维 杨凌慈缘生物技术有限公司;D-半乳糖 合肥新恩源生物技术有限公司;考马斯亮蓝蛋白测定试剂盒、MDA试剂盒、总抗氧化能力(total antioxidant capacity,T-AOC)试剂盒、总超氧化物歧化酶(total superoxide dismutase,T-SOD)试剂盒 南京建成生物研究所;其他试剂均为国产分析纯。
超微粉碎机 浙江省温岭市创力药材机械厂;JRA-2数显磁力搅拌水浴锅 金坛市科杰仪器厂;Pico17低温冷冻离心机 美国Thermo Scientific公司;TG16W微量高速离心机 长沙平凡仪器仪表有限公司;WGL-125B电热鼓风干燥箱 天津市泰斯特仪器有限公司;METASH UV-5200紫外-可见分光光度计上海元析仪器有限公司。
1.3.1 饲料的设计及制备
将基础饲料打粉,以此为基础设计特征饲料并制备成粒。依据本课题组前期研究结果,并参考齐广海等[22]的方法,特征饲料及配方如下:胡豆饲料在基础饲料中添加一定量的β-胡萝卜素和4.0 g/kg大豆异黄酮;纤维饲料在基础饲料中添加一定量的大豆膳食纤维和玉米膳食纤维;元素饲料在基础饲料中添加一定量的硒、铁、锰、钴元素,硒元素以亚硒酸钠形式添加,其他元素均以硫酸盐形式添加;复合饲料在基础饲料中综合添加一定量的β-胡萝卜素、大豆异黄酮、大豆膳食纤维和玉米膳食纤维,同时添加硒、铁、锰、钴元素,大豆异黄酮添加量为1.0 g/kg。各饲料具体营养成分见表1。
表 1 基础饲料及特征饲料的营养成分
Table 1 Nutrient contents of basic diet and characteristic diets
1.3.2 动物分组及处理
84 只小鼠随机分为7 组,每组12 只,分为年轻组、衰老组、能限组、胡豆组、纤维组、元素组和复合组。参考Rehman等[23]的造模方法,除年轻组外,其余6 组每日上午颈背部皮下注射500 mg/kg D-半乳糖,年轻组注射等体积生理盐水,持续8 周。造模同时,各组开始给予相应处理,年轻组和衰老组饲喂基础饲料,不限进食;能限组饲喂基础饲料,参考刘天娥等[24]的方法,限制30%进食;胡豆组、纤维组和元素组分别饲喂胡豆饲料、纤维饲料和元素饲料,不限进食;复合组饲喂复合饲料,限制10%进食。各组小鼠自由饮水,连续饲喂8 周,每周称质量一次,每日上午记录进食量。
1.3.3 样品采集和处理
饲喂期结束后,所有小鼠禁食、禁水12 h,称体质量,麻醉后摘眼球取血,所采集的血液在4 ℃下静置,待血清析出后,在高速离心机中4 ℃、3 000 r/min离心15 min,保留上清液并分装,保存于-80 ℃下。
取血后,麻醉小鼠迅速脊椎脱臼处死,冰上取出心脏、肝脏、脾脏、肾脏和脑组织,去除脂肪和肌膜后用冰生理盐水漂洗、滤纸吸干、称质量,并迅速冻存于-80 ℃。检测当日,取出肝脏和脑组织,称质量,加入9 倍体积的生理盐水,使用匀浆器在冰水浴中匀浆,制备质量分数10%的组织匀浆,并于4 ℃下以3 000 r/min离心15 min,取上清液进行检测。
1.3.4 脏器系数和生化指标的检测
脏器系数按以下公式计算。
式中:m1为小鼠脏器质量/g;m2为小鼠体质量/g。
按试剂盒说明书,分别对血清中的MDA浓度和T-SOD活力、肝脏组织中的组织蛋白质量浓度、MDA含量和T-AOC、脑组织中的组织蛋白质量浓度、MDA含量、T-AOC和T-SOD活力进行检测。并对其结果进行综合量化评价。
本实验数据采用SPSS 19.0软件进行统计分析,结果表示为 ±s,组间比较采用单因素方差分析。P<0.05表示两组数值间差异显著,P<0.01表示两组数值间差异极显著。
±s,组间比较采用单因素方差分析。P<0.05表示两组数值间差异显著,P<0.01表示两组数值间差异极显著。
表 2 特征饮食对D-半乳糖致衰老小鼠体质量的影响
Table 2 Effects of characteristic diets on body mass of D-galactose-induced aging mice

注:*.与年轻组比较,差异显著(P<0.05);与衰老组比较,#.差异显著(P<0.05),##.差异极显著(P<0.01);下同。
小鼠体质量的增长是反映小鼠生长发育情况的重要指标[25]。由表2可知,实验开始时各组小鼠的体质量为(36±2)g,各组之间没有显著差异(P>0.05)。实验结束时,与年轻组相比,衰老组、纤维组、胡豆组、元素组和复合组小鼠体质量差异不显著(P>0.05);与衰老组相比,能限组和胡豆组小鼠的体质量显著降低(P<0.01,P<0.05)。在体质量总增长量上,衰老组和年轻组小鼠间差异不显著(P>0.05),表明长期注射D-半乳糖对小鼠体质量的增长无显著影响;与年轻组和衰老组相比,能限组小鼠体质量均显著降低(P<0.05,P<0.01),说明能量限制能显著抑制体质量的增长;其他特征饮食组的体质量增长差异不显著。Lemon等[26]的研究证明,给予转基因生长激素小鼠一定剂量的复合膳食补充剂不会对小鼠的体质量增长造成不良影响。本研究发现特征饮食的饲喂也未对D-半乳糖致衰老小鼠的体质量增长产生显著影响,说明特征饮食对D-半乳糖致衰老小鼠的生长发育无显著的不良作用。
表 3 特征饮食对D-半乳糖致衰老小鼠脏器系数的影响
Table 3 Effects of characteristic diets on organ coef ficients of D-galactose-induced aging mice
%
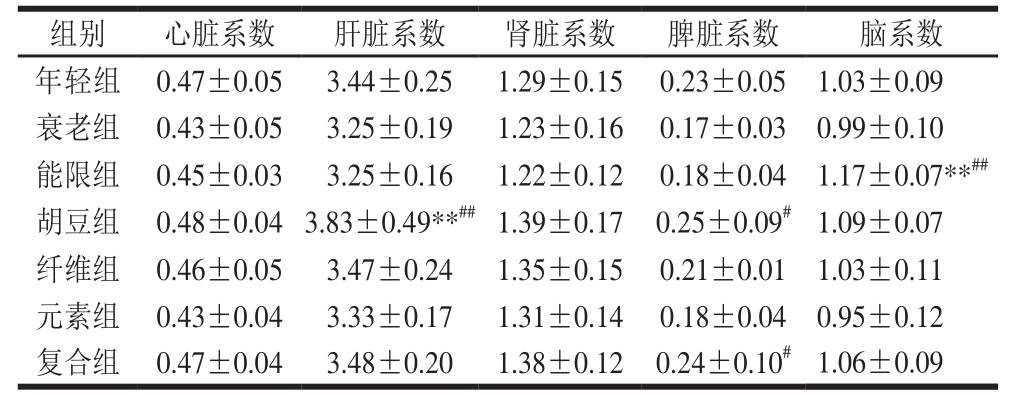
注:**.与年轻组比较差异极显著(P<0.01),下同。
实验动物的脏器系数是动物主要的生物学特性之一,能反映动物的整体营养情况以及器官病变程度,而脑、肝脏、肾脏等重要脏器的萎缩也是衰老的重要宏观表现之一[27]。由表3可知,与年轻组相比,衰老组小鼠各项脏器系数均有所下降,但差异不显著(P>0.05)。与衰老组相比,特征饮食各组的心脏系数、肝脏系数、肾脏系数、脾脏系数和脑系数均有一定程度提高,尤其是能限组小鼠的脑系数极显著升高了18.18%(P<0.01);胡豆组小鼠的肝脏系数极显著升高了17.85%(P<0.01),脾脏系数显著升高了47.06%(P<0.05);复合组小鼠的脾脏系数显著升高了41.17%(P<0.05)。王驰等[28]的研究发现,灌胃富硒玉米肽能显著提高老龄小鼠的胸腺指数,这可能与机体抗氧化酶活力提高有关。而本研究结果也表明,5 种特征饮食均有延缓衰老小鼠脏器退化的作用,其机理可能与清除自由基、提高抗氧化酶活力有关。
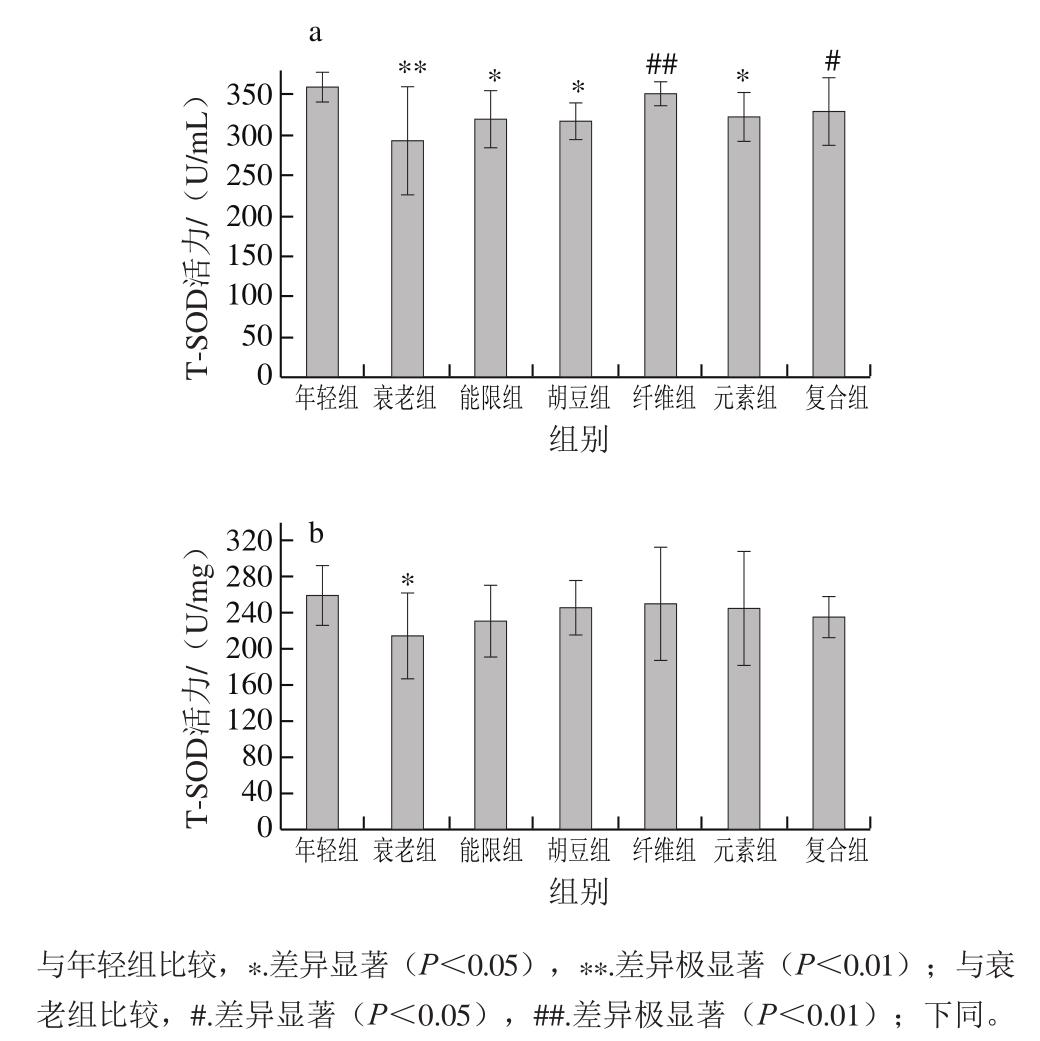
图 1 特征饮食对D-半乳糖致衰老小鼠血清(a)和脑组织(b)中T-SOD活力的影响
Fig. 1 Effects of characteristic diets on T-SOD activity in serum (a)and brain (b) of D-galactose-induced aging mice
由图1可知,与年轻组相比,衰老组小鼠在长期注射D-半乳糖的作用下,血清和脑组织中T-SOD活力分别降低了18.90%(P<0.01)和17.15%(P<0.05),表明衰老小鼠模型建立成功。经饮食干预以后,与衰老组相比,特征饮食各组小鼠血清和脑组织中T-SOD活力均有一定程度的提高,尤其是纤维组和复合组小鼠血清中的T-SOD活力分别提高了19.98%(P<0.01)和12.33%(P<0.05)。自由基是生物体氧化磷酸化过程的副产物,机体的衰老与自由基在机体内的动态平衡紧密相关[29]。自由基的清除依赖于机体防御体系抗氧化能力的强弱,SOD是机体内极为重要的自由基清除剂和抗氧化酶,能催化超氧化物通过歧化反应转化为氧气和过氧化氢,清除氧自由基从而保护细胞[30-31]。研究表明,T-SOD活力可以反映机体清除自由基的能力,是评价实验动物衰老的重要指标之一[32-33]。叶静等[34]的研究发现灌胃200 mg/kg燕麦膳食纤维能使衰老小鼠血清T-SOD提高12.19%,稍低于本研究中复合组的效果,表明融合了多种巴马长寿老人饮食营养特征的复合饮食对D-半乳糖诱导的氧化损伤有良好的作用效果,能明显减缓机体内的氧化损伤程度。
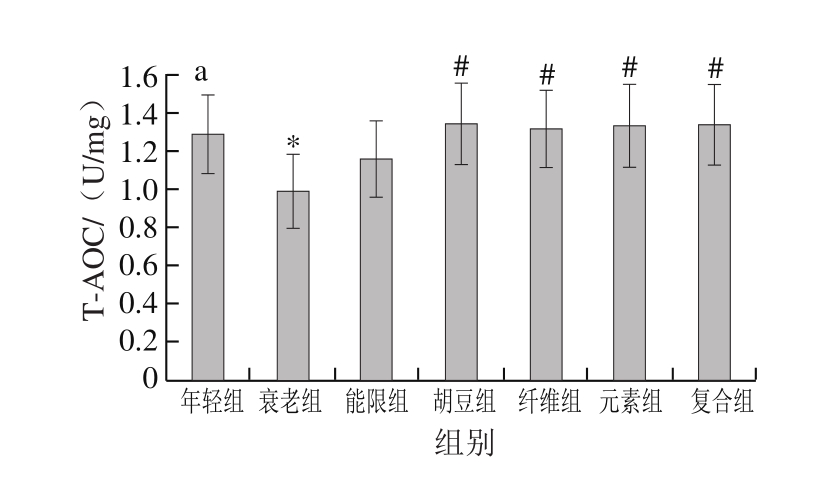
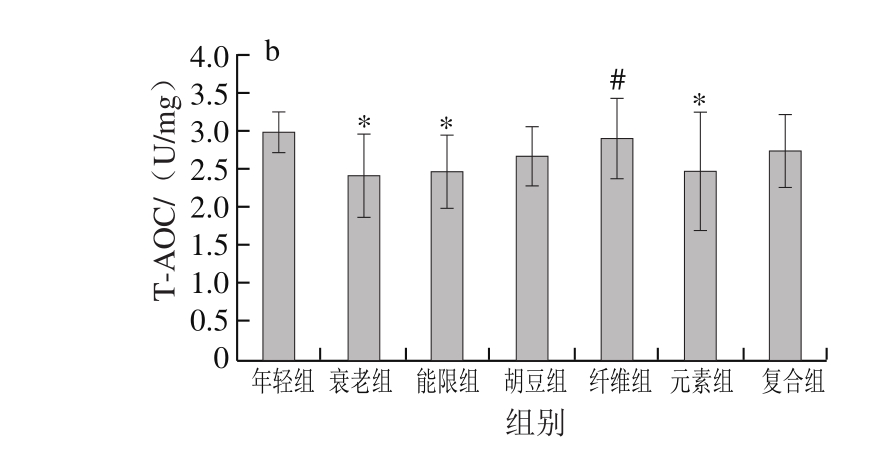
图 2 特征饮食对D-半乳糖致衰老小鼠肝脏(a)和脑组织(b)中T-AOC的影响
Fig. 2 Effects of characteristic diets on T-AOC activity in liver (a) and brain (b) of D-galactose-induced aging mice
图2显示,饮食干预以后,与年轻组相比,衰老组小鼠肝脏和脑组织中T-AOC分别降低了23.36%(P<0.05)和18.30%(P<0.05),说明D-半乳糖致衰老小鼠的抗氧化能力遭到破坏。与衰老组相比,胡豆组、纤维组、元素组和复合组小鼠肝脏的T-AOC分别显著提高了36.36%、33.33%、35.35%和35.35%(P<0.05),特征饮食各组小鼠脑组织的T-AOC也都有一定程度的提高,纤维组小鼠脑组织的T-AOC提高了19.50%(P<0.05)。
机体的T-AOC包括酶促体系和非酶促体系,是直接反映抗氧化水平的重要指标[35]。T-AOC的降低是衰老过程中重要的生理学变化,与机体的健康密切相关[36]。徐红艳等[37]的研究也表明,D-半乳糖衰老模型造模成功的标志之一是T-AOC和抗氧化酶活力的显著降低。此外,有研究表明维生素、大豆异黄酮和膳食纤维的摄入对提高衰老小鼠机体T-AOC有重要作用[38-40]。王艳红等[41]的研究表明,灌胃衰老小鼠75 mg/kg马栗提取物能使小鼠肝脏T-AOC提高17.6%,低于本研究中复合组的效果(35.35%),表明结合多种巴马长寿老人饮食营养特征的复合饮食能有效预防由氧化应激导致的氧化损伤。
表 4 特征饮食对D-半乳糖致衰老小鼠血清、肝脏和脑组织中MDA水平的影响
Table 4 Effects of characteristic diets on MDA level in serum, liver and brain of D-galactose-induced aging mice

在T-SOD活力和T-AOC的研究基础上,进一步探究了特征饮食对D-半乳糖致衰老小鼠血清、肝脏和脑组织中MDA水平的影响。从表4结果可知,经过饮食干预以后,与年轻组相比,衰老组小鼠血清、肝脏和脑组织中MDA水平分别升高了37.48%(P<0.01)、64.41%和57.02%(P<0.05),表明D-半乳糖致衰老模型小鼠体内的脂质过氧化加剧。与衰老组相比,特征饮食各组小鼠血清、肝脏和脑组织中MDA水平均有一定程度的降低,其中能限组小鼠血清和脑组织MDA水平分别降低了19.50%(P<0.05)和45.79%(P<0.01),胡豆组和元素组小鼠脑组织MDA水平分别降低了37.89%(P<0.01)和32.10%(P<0.05),纤维组小鼠血清和脑组织MDA水平分别降低了20.15%(P<0.05)和38.42%(P<0.01),复合组小鼠血清、肝脏和脑组织MDA水平分别降低了19.40%、39.18%和35.26%(P<0.05)。
MDA是脂质过氧化的产物,机体产生的自由基会进攻脂质进而导致脂质过氧化产生MDA,因此MDA含量能直接反映机体脂质过氧化水平[42]。孙荣花等[43]的研究发现,连续8 周皮下注射0.5 g/kg D-半乳糖能使小鼠整体表现出衰老症状,包括脑组织MDA含量的提高。罗磊等[44]的研究发现,绿豆膳食纤维的摄入能使衰老小鼠肝脏MDA含量降低26.00%,低于本研究中复合组的效果(39.18%),表明结合多种巴马长寿老人饮食营养特征的复合饮食能有效抑制脂质过氧化水平,维持机体健康。目前鲜有巴马长寿饮食在小鼠模型中作用的报道,而本实验以D-半乳糖致衰老模型研究并验证了巴马长寿饮食模式在抗氧化应激方面的作用,有助于解析长寿地区的饮食模式和延缓衰老间的联系。
对上述5 种饮食模式参考李全阳等[45]的方法进行综合量化评价,以便更加直观地反映不同模式的抗衰老效果,结果见表5。
表 5 5 种饮食模式综合量化评价结果
Table 5 Comprehensive evaluation of five dietary patterns

由表5可知,5 种模式的抗衰老效果排序为:复合组>胡豆组>能限组>纤维组>元素组。韩雪等[46]的研究发现,与单一抗氧化剂相比,复合抗氧化剂对小鼠清除自由基有更好的效果,这可能与抗氧化剂间的协同作用相关;Lemon等[26]的研究也表明,复合多种营养成分的膳食补充剂能更好地延缓老龄小鼠的衰老。本研究发现,复合组比其他4 种特征饮食组效果更好,说明这几种特征营养素可能存在协同作用。巴马长寿饮食模式对D-半乳糖诱导的衰老小鼠机体有着显著抗氧化作用,具有一定的开发潜力。
本研究以巴马长寿老人饮食的特征营养素为基础,设计5 种特征饮食模式,以D-半乳糖致衰老小鼠模型进行验证。结果表明,5 种特征饮食对D-半乳糖致衰老小鼠的生长发育均无不良影响,对小鼠血清、肝脏和脑组织的氧化应激都有一定抗氧化作用。对比其余4 种特征饮食,结合多种巴马长寿老人饮食营养特征的复合组饮食对衰老小鼠的血清、肝脏和脑组织MDA蓄积均有抑制作用,对肝脏MDA蓄积的抑制程度最强,能显著提高肝脏
T-AOC和血清T-SOD活力,对机体的抗氧化应激作用效果最好。对5 种模式进行综合量化评价,抗衰老效果排序为:复合组>胡豆组>能限组>纤维组>元素组。巴马长寿老人饮食模式对机体抗氧化应激有着积极作用,可能有助于机体健康,具有良好的开发潜力。
参考文献:
[1] HARMAN D. Aging: a theory based on free radical and radiation chemistry[J]. Journal of Gerontology, 1956, 11(3): 298-300.DOI:10.1093/geronj/11.3.298.
[2] ARAI Y, WATANABE S, KIMIRA M, et al. Dietary intakes of fl avonols, fl avones and isof l avones by Japanese women and the inverse correlation between quercetin intake and plasma LDL cholesterol concentration[J]. Journal of Nutrition, 2000, 130(9): 2243-2250.
[3] FINKEL T, HOLBROOK N J. Oxidants, oxidative stress and the biology of ageing[J]. Nature, 2000, 408: 239-247.DOI:10.1038/35041687.
[4] 俞卓伟, 保志军, 阮清伟, 等. 氧化应激-炎症-衰老及其与ApoE基因相关性研究进展[J]. 生理学报, 2013, 65(3): 338-346. DOI:10.13294/j.aps.2013.03.005.
[5] 林映雪, 庄朋伟, 张金保, 等. D-半乳糖剂量及小鼠性别对衰老模型的影响[J]. 天津中医药大学学报, 2013, 32(3): 144-147.DOI:10.11656/j.issn.1673-9043.2013.03.06.
[6] MICÓ V, BERNINCHES L, TAPIA J, et al. NutrimiRAging:micromanaging nutrient sensing pathways through nutrition to promote healthy aging[J]. International Journal of Molecular Sciences, 2017,18(5): 915-935. DOI:10.3390/ijms18050915.
[7] MORABIA A, ABEL T. The WHO report “preventing chronic diseases: a vital investment” and us[J]. Sozial-und Präventivmedizin SPM, 2006, 51(2): 74. DOI:10.1007/s00038-005-0015-7.
[8] Centers for Disease Control and Prevention. The power of prevention;chronic disease... the public health challenge of the 21stcentury[EB/OL].[2017-08-06]. https://stacks.cdc.gov/view/cdc/5509.
[9] CHRYSOHOOU C, STEFANADIS C. Longevity and diet. myth or pragmatism?[J]. Maturitas, 2013, 76(4): 303-307. DOI:10.1016/j.maturitas.2013.09.014.
[10] MITROU P N, KIPNIS V, THIÉBAUT A C, et al. Mediterranean dietary pattern and prediction of all-cause mortality in a US population:results from the NIH-AARP diet and health study[J]. Archives of Internal Medicine, 2007, 167(22): 2461-2468. DOI:10.1001/archinte.167.22.2461.
[11] MILLER K N, BURHANS M S, CLARK J P, et al. Aging and caloric restriction impact adipose tissue, adiponectin, and circulating lipids[J].Aging Cell, 2017, 16(3): 497-507. DOI:10.1111/acel.12575.
[12] SIMPSON S J, LE COUTEUR D G, RAUBENHEIMER D, et al.Dietary protein, aging and nutritional geometry[J]. Ageing Research Reviews, 2017, 39: 78-86. DOI:10.1016/j.arr.2017.03.001.
[13] WILLCOX D C, SCAPAGNINI G, WILLCOX B J. Healthy aging diets other than the mediterranean: a focus on the Okinawan diet[J].Mechanisms of Ageing and Development, 2014, 137(3): 148-162.DOI:10.1016/j.mad.2014.01.002.
[14] WILLCOX D C, WILLCOX B J, TODORIKI H, et al. Caloric restriction and human longevity: what can we learn from the Okinawans?[J].Biogerontology, 2006, 7(3): 173-177. DOI:10.1007/s10522-006-9008-z.
[15] 中国国家统计局. 第六次全国人口普查数据[R]. 北京: 中国国家统计局, 2010.
[16] LÜ J, WANG W, KRAFFT T, et al. Effects of several environmental factors on longevity and health of the human population of Zhongxiang, Hubei, China[J]. Biological Trace Element Research,2011, 143(2): 702-716. DOI:10.1007/s12011-010-8914-8.
[17] WANG F, YU T, HUANG G, et al. Gut microbiota community and its assembly associated with age and diet in Chinese centenarians[J].Journal of Microbiology and Biotechnology, 2015, 25(8): 1195-1204.DOI:10.4014/jmb.1410.10014.
[18] WANG F, HUANG G, CAI D, et al. Qualitative and semiquantitative analysis of fecal bifidobacterium species in centenarians living in Bama, Guangxi, China[J]. Current Microbiology, 2015, 71(1): 143-149. DOI:10.1007/s00284-015-0804-z.
[19] 蔡达, 伍国杰, 黄国宏, 等. 广西巴马80 岁以上老人食物及营养素摄入特征研究[J]. 食品工业科技, 2015, 36(21): 358-363; 380.DOI:10.13386/j.issn1002-0306.2015.21.066.
[20] CAI D, ZHAO S, LI D, et al. Nutrient intake is associated with longevity characterization by metabolites and element prof i les of healthy centenarians[J]. Nutrients, 2016, 8(9): 1-19. DOI:10.3390/nu8090564.
[21] CAI D, LI D, ZHAO S, et al. A correlation between diet and longevity characterization by means of element prof i les in healthy people over 80 years from a Chinese longevous region[J]. Biological Trace Element Research, 2015, 165(1): 18-29. DOI:10.1007/s12011-015-0233-7.
[22] 齐广海, 武书庚. 安全高效预混合饲料配制技术[M]. 北京: 化学工业出版社, 2012: 1-310.
[23] REHMAN S U, SHAH S A, ALI T, et al. Anthocyanins reversed D-galactose-induced oxidative stress and neuroinf l ammation mediated cognitive impairment in adult rats[J]. Molecular Neurobiology, 2017,54(1): 255-271. DOI:10.1007/s12035-015-9604-5.
[24] 刘天娥, 赵健亚, 陈刚, 等. 白藜芦醇和能量限制对大鼠肝脂肪变性影响[J]. 中国公共卫生, 2014, 30(7): 915-917. DOI:10.11847/zgggws2014-30-07-20.
[25] 王喆, 陆承荣, 罗渊, 等. 注射D-半乳糖致大鼠衰老效应的评价[J]. 中国医药导报, 2013, 10(9): 25-26; 36. DOI:10.3969/j.issn.1673-7210.2013.09.010.
[26] LEMON J A, AKSENOV V, SAMIGULLINA R, et al. A multiingredient dietary supplement abolishes large-scale brain cell loss,improves sensory function, and prevents neuronal atrophy in aging mice[J]. Environmental and Molecular Mutagenesis, 2016, 57(5): 382-404. DOI:10.1002/em.22019.
[27] 罗磊, 张冰洁, 关宁宁, 等. 金银花叶黄酮对衰老模型小鼠的体内抗氧化作用[J]. 食品科学, 2017, 38(19): 171-176. DOI:10.7506/spkx1002-6630-201719027.
[28] 王驰, 张久亮, 尧咏, 等. 富硒玉米蛋白水解物抗氧化活性研究[J]. 中国粮油学报, 2017, 32(2): 49-56. DOI:10.3969/j.issn.1003-0174.2017.02.009.
[29] 夏世金, 孙涛, 吴俊珍. 自由基、炎症与衰老[J]. 实用老年医学,2014, 28(2): 100-103.
[30] 董亮, 何永志, 王远亮, 等. 超氧化物歧化酶(SOD)的应用研究进展[J]. 中国农业科技导报, 2013, 15(5): 53-58. DOI:10.3969/j.issn.1008-0864.2013.05.08.
[31] 钟灵, 王振富, 文德鉴. 黄芪多糖抗衰老作用的实验研究[J]. 中国应用生理学杂志, 2013, 29(4): 350-352. DOI:10.13459/j.cnki.cjap.2013.04.007.
[32] 孙玉军, 江昌俊, 任四海. 秀珍菇多糖对D-半乳糖致衰老小鼠的保护作用[J]. 食品科学, 2017, 38(5): 251-256. DOI:10.7506/spkx1002-6630-201705041.
[33] 蒲秀瑛, 李海兵, 李晓玥, 等. 归芪多糖对衰老大鼠肾组织氧化损伤作用的研究[J]. 中医药学报, 2017, 45(2): 15-18. DOI:10.3969/j.issn.1002-2392.2017.02.004.
[34] 叶静, 肖美添, 刘青, 等. 江蓠藻膳食纤维对衰老小鼠免疫功能与抗氧化作用的影响[J]. 食品工业科技, 2009, 30(11): 287-290.DOI:10.13386/j.issn1002-0306.2009.11.086.
[35] 邓洋, 王艳茹, 林艳伟. 苦荞茶对D-半乳糖所致衰老小鼠T-AOC和MAO影响的实验研究[J]. 食品科技, 2014, 23(5): 76-78.DOI:10.13684/j.cnki.spkj.2014.05.019.
[36] 邓洋, 张利霞. 苦荞茶对D-半乳糖所致衰老小鼠MDA和MAO影响的实验研究[J]. 食品科技, 2014, 23(5): 678-680. DOI:10.13684/j.cnki.spkj.2014.05.019.
[37] 徐红艳, 刘富国, 于阳阳, 等. 东北山核桃仁油对D-半乳糖衰老小鼠抗氧化能力的影响[J]. 食品科学, 2012, 33(7): 266-269.
[38] 余桂荣, 张志萍. 饮食控制与延缓衰老[J]. 山东医学高等专科学校学报, 2015, 37(6): 406-406. DOI:10.3969/j.issn.1674-0947.2015.06.003.
[39] 彭辉, 王欢. 膳食纤维与老年健康[J]. 中国老年学杂志, 2014,34(22): 6510-6512. DOI:10.3969/j.issn.1005-9202.2014.22.132.
[40] 韩瑞敏, 程红, 李丹丹, 等. 类胡萝卜素的抗氧化自由基分子反应机理研究[C]//中国化学会学术年会. 北京: 中国化学会, 2014: 28.
[41] 王艳红, 高江霞, 孟敏, 等. 马栗籽提取物对D-半乳糖所致小鼠衰老模型的影响[J]. 中国新药杂志, 2017, 26(2): 208-213.
[42] 崔玮, 韩春, 赵巧华, 等. 苹果皮总黄酮对D-半乳糖致衰老模型小鼠抗氧化能力的影响[J]. 食品科学, 2014, 35(15): 258-262.DOI:10.7506/spkx1002-6630-201415052.
[43] 孙荣花, 李爽, 邵莹, 等. D-半乳糖致小鼠亚急性衰老模型方法优化[J]. 中药药理与临床, 2015(4): 293-297. DOI:10.13412/j.cnki.zyyl.2015.04.104.
[44] 罗磊, 王雅琪, 马丽苹, 等. 绿豆皮可溶性膳食纤维的抗氧化作用[J]. 食品科学, 2018, 39(3): 182-187. DOI:10.7506/spkx1002-6630-201803028.
[45] 李全阳, 夏文水. 酸乳质量综合评价体系探讨[J]. 中国乳品工业,2003, 31(6): 29-33. DOI:10.3969/j.issn.1001-2230.2003.06.008.
[46] 韩雪, 陈小连, 徐建雄. 复合抗氧化剂对小鼠氧化应激与自由基代谢的影响[J]. 上海交通大学学报(农业科学版), 2010, 28(6): 526-529. DOI:10.3969/j.issn.1671-9964.2010.06.009.
Effect of Bama Longevity Dietary Patterns on Antioxidant Stress in a Mouse Model of Aging
SONG Qi1, AI Lianzhong2, LU Hongyan1, HAO Dan1, CHEN Yu1, SUN Hui1, LAN Haijing1, ZHANG Zutao3, LI Quanyang1,*
(1. College of Light Industry and Food Engineering, Guangxi University, Nanning 530004, China;2. School of Medical Instrument and Food Engineering, University of Shanghai for Science and Technology, Shanghai 200093, China;3. Shibu of Guangxi Dairy Co. Ltd., Nanning 530021, China)
Abstract:In order to validate the anti-aging effect of Bama longevity dietary patterns, a mouse model of aging was established by subcutaneously injecting D-galactose (500 mg/(kg·d)) into animals once daily for eight consecutive weeks.Meanwhile, the representative nutrients in Bama longevity dietary patterns (including calorie restriction and intake of VA and soybean isof l avones, dietary fiber and trace elements such as iron, manganese, cobalt and selenium) were selected to design five characteristic diets, calorie restriction, VA and soybean isof l avones, dietary fiber, trace elements and compound diet, for feeding aging mice. The contents of malondialdehyde (MDA) in serum, liver and brain, the activity of total superoxide dismutase (T-SOD) in serum and brain, and total antioxidant capacity (T-AOC) in liver and brain were measured.The results showed that all five diets could inhibit the accumulation of MDA in aging mice, while the activity of antioxidant enzymes and T-AOC were improved when compared with the aging model group. Through positive processing and nondimensionalization, it was found that compound nutrition had the best anti-oxidative stress effect , which could inhibit the accumulation of MDA in serum, liver and brain (P < 0.05), increase the activity of T-SOD by 12.33% (P < 0.05), and signif i cantly increase the activity of T-AOC in liver by 35.35% (P < 0.05). A comprehensive evaluation showed that the antiaging effect of 5 diets followed the decreasing order of compound diet > VA and soybean isof l avones > calorie restriction> dietary fiber > trace elements. These results demonstrate that the Bama longevity diet pattern has a signif i cant protective effect against oxidative stress in D-galactose-induced aging mice, indicating its great development potential.
Keywords:Bama longevity dietary patterns; anti-oxidative stress effect; D-galactose; aging model
SONG Qi, AI Lianzhong, LU Hongyan, et al. Effect of Bama longevity dietary patterns on antioxidant stress in a mouse model of aging[J]. Food Science, 2018, 39(19): 147-153. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201819023. http://www.spkx.net.cn
宋奇, 艾连中, 鲁红岩, 等. 巴马长寿饮食模式在衰老小鼠模型中的抗氧化应激效果[J]. 食品科学, 2018, 39(19):147-153. DOI:10.7506/spkx1002-6630-201819023. http://www.spkx.net.cn
引文格式:
文章编号:1002-6630(2018)19-0147-07
文献标志码:A
中图分类号:R154.1
DOI:10.7506/spkx1002-6630-201819023
*通信作者简介:李全阳(1964—),男,教授,博士,研究方向为营养健康与功能性食品。E-mail:liquanyang@gxu.edu.cn
第一作者简介:宋奇(1993—),男,硕士研究生,研究方向为食品营养与健康长寿。E-mail:1017380941@qq.com
基金项目:国家自然科学基金面上项目(31371762)
收稿日期:2017-08-06