副干酪乳杆菌FM-M9-1对D-半乳糖诱导氧化损伤小鼠的修复机制
王 英,夏秀东,董 月,刘小莉,周剑忠*
(江苏省农业科学研究院农产品加工研究所,江苏 南京 210014)
摘 要:目的:分析副干酪乳杆菌FM-M9-1修复氧化损伤的作用机制。方法:利用D-半乳糖诱导氧化损伤模型小鼠,以高、低剂量((5.01±0.05)×1010、(5.01±0.05)×108CFU/mL)FM-M9-1活菌体灌胃小鼠,以VC作为阳性药物对照,60 d后测定小鼠的肾和肝组织中超氧化物歧化酶(superoxide dismutase,SOD)、谷胱甘肽过氧化物酶(glutathione peroxidase,GSH-Px)活力及丙二醛、蛋白质羰基含量,同时利用荧光实时定量聚合酶链式反应法分析小鼠肝脏和肾脏组织Sod、Gsh-Px、Nrf2和Trx基因的表达水平。结果:FM-M9-1能不同程度地提高小鼠肝和肾组织中的GSH-Px和SOD活力,降低蛋白质羰基和丙二醛含量,提高与抗氧化功能相关的Sod、Gsh-Px、Nrf2和Trx基因的表达水平。结论:FM-M9-1上调小鼠体内抗氧化相关基因Sod、Gsh-Px、Nrf2和Trx的表达,提高抗氧化酶SOD、GSH-Px活力,抑制体内的蛋白质和脂肪的氧化损伤,最终达到修复小鼠体内氧化损伤的目的。
关键词:副干酪乳杆菌;D-半乳糖;小鼠;氧化损伤;修复机制
氧化应激是生物体内的活性氧/氮的产生数量超过体内抗氧化保护机制清除活性氧/氮数量的一种状态[1]。在氧化应激状态下形成的活性氧(reactive oxygen species,ROS)如超氧阴离子自由基、羟自由基、过氧化氢,能针对性地破坏生物膜脂质磷脂中的不饱和脂肪酸,导致细胞的系列过氧化反应,形成的氧化产物进一步作用于细胞的核酸和蛋白质,破坏细胞的正常功能,从而产生不同程度的氧化损伤[2-7]。目前研究结果显示,与氧化应激有关的疾病涉及到机体系统的各个方面,包括阿尔茨海默病、帕金森病、肌萎缩性脊髓侧索硬化症、哮喘、过敏、糖尿病等疾病,另外还有些高血压、高血脂、冠心病和癌症等都与氧化损伤有着十分密切的关系[8-10]。
大量关于乳酸菌体内、外的抗氧化功能研究结果显示,乳酸菌细胞和无细胞提取物在体外具有清除自由基、抑制脂质过氧化、螯合金属离子等活性[11-17]。体内摄入乳酸菌对机体或细胞的氧化应激具有显著的调节作用,能提高体内抗氧化相关功能基因的表达并降低体内过氧化产物的活力[18-20]。
目前,乳酸菌抗氧化功能一般都是通过体外筛选获得,通过体外相关抗氧化指标的测定筛选乳酸菌[21-23]。但是体外筛选的环境与体内环境相差很大,已有很多学者对此提出异议,认为乳酸菌缓解氧化应激能力必须通过体内模型的验证才有研究意义和应用价值。目前比较常用的一种小鼠氧化损伤模型是D-半乳糖长期给药造成小鼠衰老的模型,这个模型比较接近小鼠自然衰老,表现出小鼠寿命缩短、体内氧化压力上升等,可以用来观察药物的抗氧化作用。
副干酪乳杆菌FM-M9-1来源于传统自然发酵食品,是具有优良的体外抗氧化能力的乳酸菌[24]。本研究以D-半乳糖诱导的氧化应激损伤模型小鼠为研究对象,研究饲喂副干酪乳杆菌FM-M9-1对氧化应激的体内干预和缓解作用,进而揭示缓解氧化损伤的机制,为开发针对氧化还原失衡和氧化应激损伤的功能性产品提供可靠的、具有自主知识产权的乳酸菌菌株资源。
1 材料与方法
1.1 动物、材料与试剂
实验动物采用SPF级昆明种雄性小鼠,体质量(20±2)g,由扬州大学动物中心提供,许可证号:SCXK(苏)2012-0004。实验动物普通饲料由青龙山饲料物厂提供。
副干酪乳杆菌FM-M9-1由江苏省农业科学院农产品加工研究所食品生物工程研究室保存。
丙二醛(malondialdehyde,MDA)试剂盒、超氧化物歧化酶(superoxide dismutase,SOD)试剂盒、谷胱甘肽与谷胱甘肽过氧化物酶(glutathione peroxidase,GSH-Px)试剂盒 南京建成生物工程研究所;TRIzol总RNA提取试剂盒、反转录试剂盒、荧光定量试剂盒 大连宝生物科技公司;引物由上海生物工程有限公司合成,其他常规试剂均为市售分析纯。
1.2 仪器与设备
ABI7500Fast实时荧光定量聚合酶链式反应(real-time quantitative polymerase chain reaction,qPCR)仪 美国应用生物系统公司;UV-1600PC紫外分光光度计 上海美普达科技有限公司;YQX-11厌氧培养箱 上海跃进医疗器械有限公司;SW-CJ-1C型双人单面净化工作台 苏州净化设备有限公司;3K15台式冷冻离心机德国Sigma公司。
1.3 方法
1.3.1 小鼠饲养
小鼠适应一周后,称体质量,然后随机分成5 组,每组10 只。将小鼠同室分笼喂养,采用自然光照,自由采食和饮水。饲养环境温度(25±2)℃,相对湿度60%,实验周期为60 d。
1.3.2 菌株FM-M9-1培养及菌体处理
挑取FM-M9-1单菌落至MRS液体试管中,37 ℃静止培养18~20 h至稳定初期,以此为接种液,按体积分数3%的接种量转接到MRS液体培养基中培养18 h,上述培养好的菌液5 000 r/min离心5 min收集菌体,用生理盐水洗2 次后重悬菌体,调整菌体浓度分别为(5.01±0.05)×108CFU/mL和(5.01±0.05)×1010CFU/mL,备用。
1.3.3 小鼠分组
对照组:颈背部皮下每天注射200 mg/(kg·d)生理盐水,灌胃0.2 mL生理盐水;模型组、实验组和VC组颈背部皮下每天注射D-半乳糖(200 mg/(kg·d))。其中,模型组灌胃0.2 mL生理盐水;实验组中,低剂量组灌胃0.2 mL菌体浓度为(5.01±0.05)×108CFU/mL的FM-M9-1菌液;高剂量组灌胃0.2 mL菌体浓度为(5.01±0.05)×1010CFU/mL的FM-M9-1菌液;VC组灌胃200 mg/(kg·d) VC。
1.3.4 小鼠肝脏和肾组织样品的处理
实验结束后,将小鼠禁食12~18 h,断颈处死,迅速取出肝脏和肾脏,置于冰冷生理盐水中漂洗拭干,称质量,制备肾脏和肝组织匀浆液用于氧化应激标志物的测定和分析。
肝组织匀浆的制备:每只取出小鼠肝组织总质量的10%,加入9 倍组织质量的预冷生理盐水,充分研磨匀浆,3 000 r/min离心15 min,弃沉淀,所得上清液即为肝匀浆液,备用。肾组织匀浆的制备同肝组织。
肝、肾组织匀浆液中蛋白质量浓度采用考马斯亮蓝法测定,以牛血清白蛋白为标准蛋白质,制作标准曲线,得到蛋白质量浓度(x/(mg/mL))与OD595nm(y)的回归方程为y=0.010 9x+0.081 4(R2=0.998 6)。
1.3.5 氧化应激标志物的检测与分析
利用试剂盒测定小鼠肝和肾脏组织中的GSH-Px、SOD活力以及蛋白质羰基、MDA含量。
1.3.6 qPCR法检测Sod、Gsh-Px、Nrf2和Trx基因的相对表达水平
用TRIzol试剂提取各实验组小鼠肝和肾脏组织的RNA,超微量核酸分析仪测定浓度,按照逆转录试剂盒说明书进行反转录合成cDNA,SYBR Select Mast Mix:ABI配制10 µL反应体系,采用ABI7500Fast qPCR检测Sod、Gsh-Px、Nrf2和Trx基因的表达水平,引物序列如表1所示。以β-actin为内参基因,Ct值通过荧光定量仪的分析软件给出,最后基因的表达水平用2-△Ct表示。
表1 qPCR引物序列
Table1 Primer sequences used for qPCR

基因 引物序列(5’-3’) Ct值β-actin 正向:ATATCGCTGCGCTGGTCGTC 23反向:AGGATGGCGTGAGGGAGAGC Sod 正向:ACGGTGTGGCCAATGTGTC 22反向:TTACTGCGCAATCCCAATCA Gsh-Px 正向:GGTTTCCCGTGCAATCAGTTCG 23反向:GAGCGCAGTGGGGTCGTCA Trx 正向:AGCCCTTCTTCCATTCCCTCTGT 23反向:CTCCCCCACCTTTTGACCCTTTTT Nrf2 正向:CTAAAGCACAGCCAGCACATTCTC 23反向:CTGGGCGGCGACTTTATTCTTAC
1.4 数据分析
采用Microsoft Excel 2010软件进行实验数据的统计和整理,采用SPSS 15.0和One-way ANOVA软件进行统计和显著性分析,数据以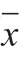 ±s表示,P<0.05表示差异显著。
±s表示,P<0.05表示差异显著。
2 结果与分析
2.1 不同处理对氧化损伤小鼠肝脏和肾组织GSH-Px活力的影响
蛋白质羰基和MDA含量、SOD、GSH-Px活力是反映机体氧化应激活力的4 个常见标志物。已有研究结果显示,乳酸菌处理氧化损伤小鼠,能不同程度地提高小鼠机体组织中SOD、GSH-Px等抗氧化酶的活力,同时降低蛋白质羰基化和MDA含量。Wang Yuzhen等[18]利用Lactobacillus casei Zhang菌株处理内毒素和D-半乳糖诱导的氧化损伤小鼠,发现Lactobacillus casei Zhang能够显著提高小鼠肝脏中的SOD活力,并显著降低MDA含量。Kamaladevi等[20]利用Lactobacillus casei处理有机磷杀虫剂诱导氧化损伤的秀丽隐杆线虫,发现Lactobacillus casei能够显著提高线虫的过氧化氢酶、SOD、GSH-Px、谷胱甘肽转移酶等相关抗氧化酶的活力,进而缓解氧化损伤。Zhang Li等[25]利用Lactobacillus plantarum C88处理过氧化氢诱导的氧化损伤Caco-2细胞,发现Lactobacillus plantarum C88能够显著降低MDA含量,显著提高细胞的SOD和总抗氧化酶活力,并呈现量效关系。
表2 不同处理对小鼠肝、肾GSH-Px活力的影响
Table2 Effects of different treatment on GSH-Px activity in liver and kidney of mice

注:模型组与对照组相比,#.差异显著(P<0.05),##.差异极显著(P<0.01);实验组、VC组与模型组相比,**.差异极显著(P<0.01),***.差异高度显著(P<0.001)。下同。
组别 肾脏GSH-Px活力/(U/mg pro)肝脏GSH-Px活力/(U/mg pro)对照组 198.14±25.80 209.41±17.77模型组 154.80±9.58#157.32±6.69##低剂量组 165.47±8.37 178.31±4.75高剂量组 238.37±8.90** 224.49±5.85**VC组 227.02±11.76** 259.65±6.03***
从表2可以看出,与对照组相比,模型组小鼠的肾脏、肝脏GSH-Px活力显著降低(P<0.05,P<0.01),表明D-半乳糖诱导的小鼠氧化损伤模型比较成功。与模型组相比,低、高剂量组均能不同程度地提高肝脏和肾脏组织中GSH-Px活力,但是低剂量组差异不显著。
2.2 不同处理对氧化损伤小鼠肝脏和肾组织SOD活力的影响
在机体内,SOD具有清除超氧阴离子自由基的功能,保护细胞免受超氧阴离子自由基造成的损伤,在氧化还原平衡与氧化应激酶调节机制中扮演着重要角色。
表3 不同处理对小鼠肝、肾SOD活力的影响
Table3 Effect of different treatments on SOD activity in liver and kidney of mice

组别 肾脏SOD活力/(U/mg pro)肝脏SOD活力/(U/mg pro)对照组 84.43±11.26 96.54±8.45模型组 54.52±4.67##79.10±3.14##低剂量 58.63±3.43 84.44±2.14高剂量 95.26±4.39*** 105.98±2.79***VC组 81.34±5.01** 101.59±2.89***
由表3可知,与模型组相比,低、高剂量FM-M9-1均能不同程度地提高肝脏和肾脏组织中SOD活力,但是低剂量组小鼠的肝脏和肾脏组织中SOD活力升高不显著(P>0.05)。
2.3 不同处理对氧化损伤小鼠肝脏和肾组织蛋白质羰基含量的影响
表4 不同处理对小鼠肝、肾蛋白质羰基含量的影响
Table4 Effects of different treatments on protein carbonyl content in liver and kidney of mice
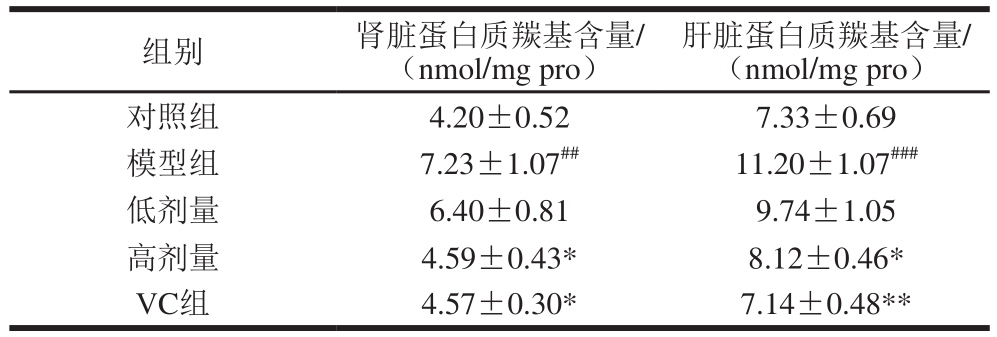
注:模型组与对照组相比,###.差异高度显著(P<0.001);实验组、VC组与模型组相比,*.差异显著(P<0.05),下同。
肝脏蛋白质羰基含量/(nmol/mg pro)对照组 4.20±0.52 7.33±0.69模型组 7.23±1.07##11.20±1.07###低剂量 6.40±0.81 9.74±1.05高剂量 4.59±0.43* 8.12±0.46*VC组 4.57±0.30* 7.14±0.48**组别 肾脏蛋白质羰基含量/(nmol/mg pro)
由表4可知,与对照组相比,模型组小鼠的肾脏和肝脏组织中蛋白质羰基的含量都显著升高(P<0.01或P<0.001),表明D-半乳糖诱导的小鼠氧化损伤模型比较成功。与模型组相比,不同剂量组均能不同程度地下调肝脏和肾脏组织中蛋白质羰基含量,低剂量组的小鼠的肝和肾中蛋白质羰基含量下降不显著(P>0.05)。
2.4 不同处理对氧化损伤小鼠肝脏和肾组织MDA含量的影响
MDA含量是反映机体内脂质过氧化程度的一项生物标志物,可以间接反映机体和细胞受ROS与自由基攻击的氧化应激程度。
表5 不同处理对小鼠肝、肾MDA含量的影响
Table5 Effects of different treatments on MDA content in liver and kidney of mice

组别 肾脏MDA含量/(nmol/mg pro)肝脏MDA含量/(nmol/mg pro)对照组 4.85±0.54 4.12±0.46模型组 8.62±1.19##7.52±1.08###低剂量 7.85±1.01 6.70±0.69*高剂量 5.76±0.49* 4.66±0.40**VC组 5.42±0.36** 4.67±0.29**
由表5可知,与模型组相比,低、高剂量组均能不同程度地下调肝脏和肾脏组织中MDA含量,低剂量组小鼠肾脏组织的MDA含量与模型组相比差异不显著(P>0.05),肝脏组织中的MDA含量与模型组相比显著降低(P<0.05)。
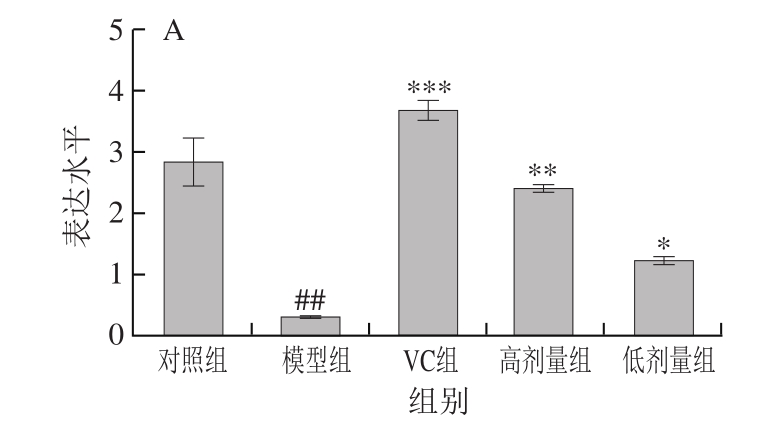
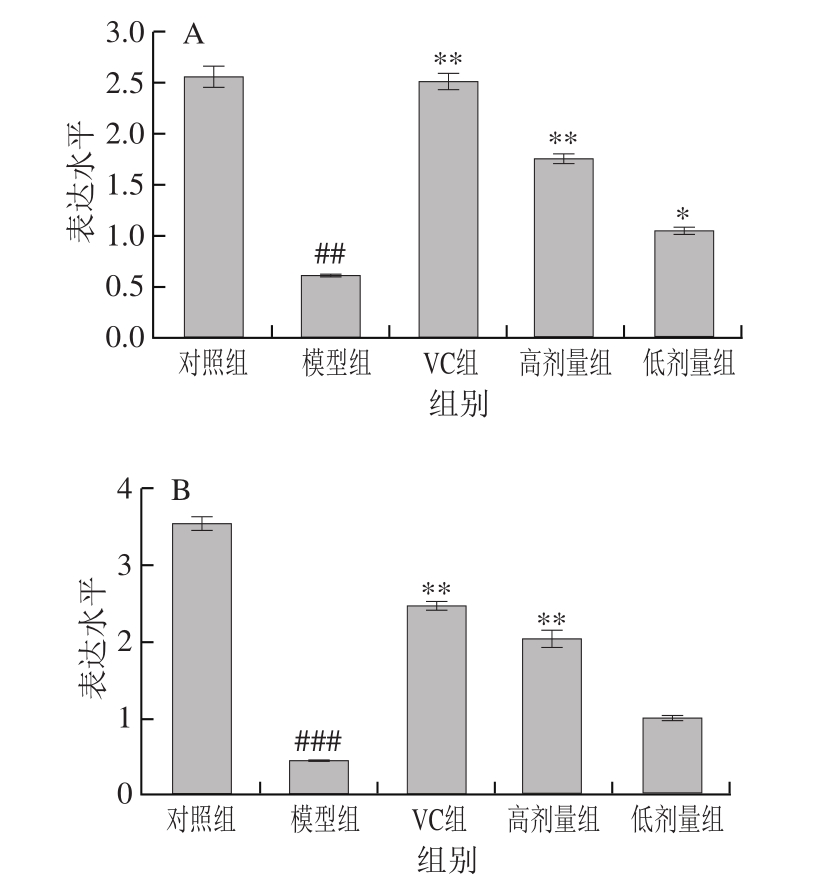
2.5 不同处理对氧化损伤小鼠肝脏和肾组织中Sod基因表达水平的影响
如图1A所示,与对照组相比,模型组小鼠肾组织Sod基因表达水平极显著降低(P<0.01)。与模型组相比,VC组和高剂量组Sod基因表达水平极显著提高(P<0.01或P<0.001),低剂量FM-M9-1能够显著提高Sod基因的表达水平(P<0.05)。图1B显示,与对照组相比,模型组小鼠肝组织中的Sod基因表达水平极显著降低(P<0.01)。与模型组相比,不同剂量乳酸菌和VC处理都能不同程度地提高Sod基因的表达水平。
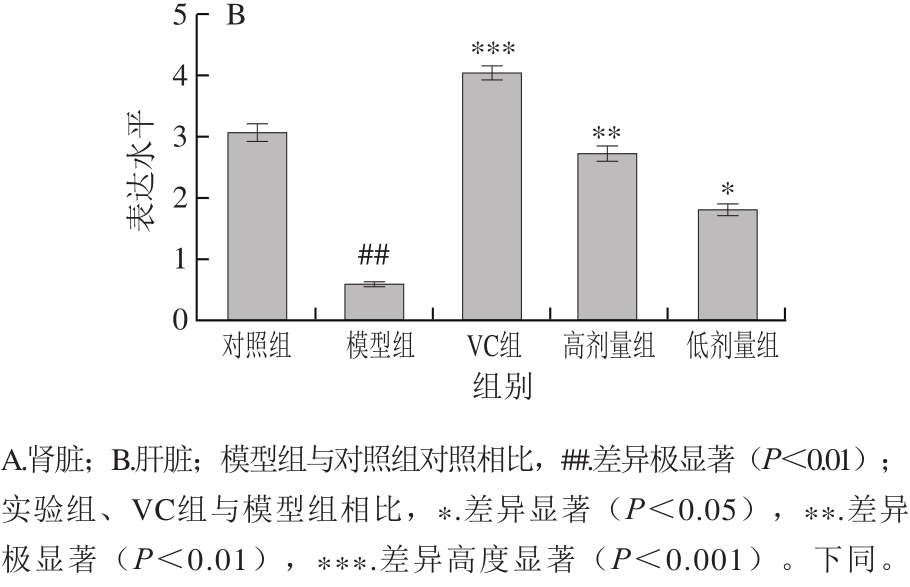
2.6 不同处理对氧化损伤小鼠肝脏和肾组织中Gsh-Px基因表达水平的影响
GSH-Px是机体抗过氧化屏障的重要组分,其表达水平的高低与机体对过氧化损伤的修复有密切关系。
由图2可知,与对照组相比,模型组小鼠肾和肝组织中的Gsh-Px基因表达水平高度显著降低(P<0.001)。与模型组相比,高剂量乳酸菌和VC处理都能不同程度地提高Gsh-Px基因的表达水平,低剂量组变化不显著。
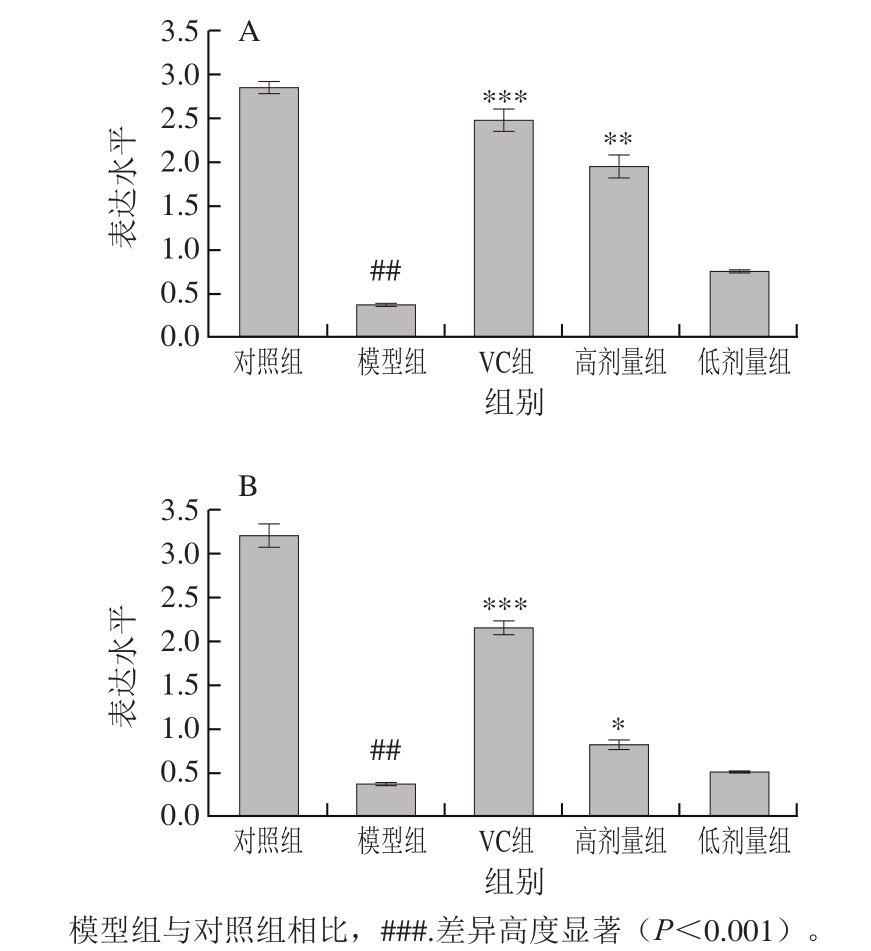
2.7 不同处理对氧化损伤小鼠肝脏和肾组织中Nrf2基因表达水平的影响
核因子E2相关因子2(nuclear factor E2-related factor-2,Nrf2)是氧化应激的防御机制之一,它调节并激活细胞内抗氧化应激系统中和活性氧、促进细胞存活、保持细胞内氧化还原稳定[26]。Zhou Ningna等[27]利用来源于Panax notoginseng的皂素处理损伤的SH-SY5Y细胞,发现Panax notoginseng的皂素能够通过上调Nrf2的表达并增强Nrf2及其下游抗氧化系统活力,主要包括血红素加氧酶和谷胱甘肽转移酶,从而缓解细胞的ROS积累。
由图3可知,与对照组相比,模型组小鼠肾和肝组织中的Nrf2基因表达水平高度显著降低(P<0.001)。与模型组相比,VC和高剂量FM-M9-1能够极显著提高肝和肾组织中Nrf2基因的表达水平(P<0.01或P<0.001)。低剂量FM-M9-1能够显著上调肾组织Nrf2基因表达水平(P<0.05),肝组织中Nrf2基因表达水平未显著上调(P>0.05)。
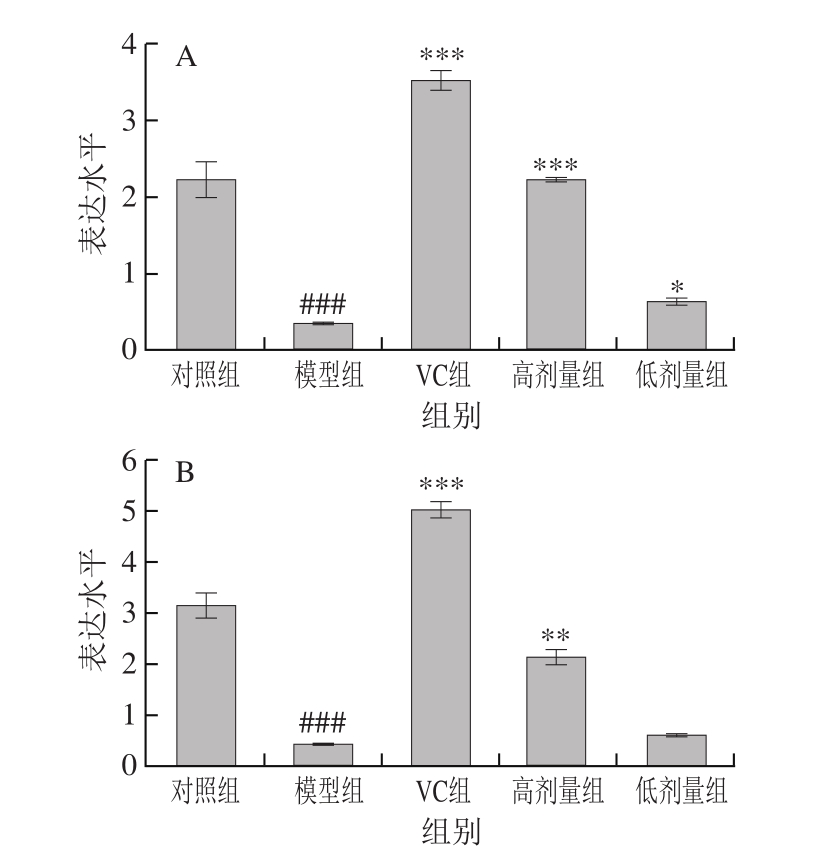
2.8 不同处理对氧化损伤小鼠肝脏和肾组织中Trx基因表达水平的影响
硫氧还蛋白(thioredoxin,Trx)、硫氧还蛋白还原酶(thioredoxin reductase,TrxR)与烟酰胺腺嘌呤二核苷磷酸(nicotinamide adenine dinucleotide phosphate,NADPH)构成的Trx系统是一个广泛分布于原核生物和真核生物的NADPH依赖的二硫化物还原酶系统,在机体氧化还原平衡及细胞增殖中发挥重要的调节作用[28],Trx能够直接清除细胞内过氧化氢及氧自由基等ROS,调节细胞内氧化还原平衡[29-30]。Yamada等[31]证实Trx能通过清除ROS保护高氧诱导的肺上皮细胞损伤。Tanaka等[32]在实验中观察到Trx缺失的细胞体内有ROS增加、凋亡细胞增多以及细胞色素c表达上调等现象,从而推断Trx能清除线粒体内的ROS并在调节线粒体细胞凋亡的信号转导过程中发挥重要作用。
由图4可知,与对照组相比,模型组小鼠肾和肝组织中Trx基因表达水平显著下调(P<0.01或P<0.001)。与模型组相比,高剂量FM-M9-1能够极显著上调Trx基因的表达水平(P<0.01);低剂量组肾组织Trx基因表达水平显著上调(P<0.05),但在肝组织中未显著上调(P>0.05)。
3 结 论
利用D-半乳糖诱导氧化损伤模型小鼠,研究不同剂量的FM-M9-1菌体对氧化损伤小鼠的修复作用,同时以VC作为阳性药物对照,测定肾和肝组织中SOD、GSH-Px、MDA、蛋白质羰基含量,同时利用qPCR方法对小鼠肝脏和肾脏组织的Sod、Gsh-Px、Nrf2和Trx基因的表达水平进行分析,揭示FM-M9-1缓解氧化损伤的作用机制。结果表明:低、高剂量FM-M9-1均能不同程度地提高小鼠肝和肾组织中的GSH-Px和SOD的活力,降低小鼠肝和肾组织中的蛋白质羰基和MDA含量,提高小鼠肝脏和肾脏组织中与抗氧化功能相关的Sod、Gsh-Px、Nrf2和Trx基因的表达水平。推测FM-M9-1通过上调小鼠体内抗氧化相关基因Sod、Gsh-Px、Nrf2和Trx的表达,提高抗氧化酶SOD、GSH-Px活力,抑制体内的蛋白质和脂肪的氧化损伤,最终达到缓解小鼠氧化损伤的作用。
参考文献:
[1] PERSSON T, POPESCU B O, CEDAZO-MINGUEZ A, et al.Oxidative stress in Alzheimer’s disease: why did antioxidant therapy fail?[J]. Oxidative Medicine Cell and Longevity, 2014, 2014: 1-11.DOI:10.1155/2014/427318.
[2] URBAN T, HURBAIN I, URBAN M, et al. Oxidants and antioxidants.biological effects and therapeutic perspectives[J]. Annales de Chirurgie, 1995, 49(5): 427-434.
[3] 相启森, 孟旭, 乔燕, 等. 鼠尾草酸对自由基诱导蛋白质氧化损伤的保护作用[J]. 食品科学, 2013, 34(15): 281-284. DOI:10.7506/spkx1002-6630-201315058.
[4] PATETSINI E, DIMITRIADIS B K, KALOYIANNI M. Exposure of mussels Mytilus galloprovinciallis, to environmental pesticides.study of LMS, ROS, DNA damage, protein carbonylation and antioxidantcapacity for their use as biomarkers[J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2012, 163: 2985-3023. DOI:10.1016/j.cbpa.2012.05.082.
[5] DARZYNKIEWICZ Z, ZHAO H, HALICKA H D, et al. In search of antiaging modalities: evaluation of mTOR- and ROS/DNA damage-signaling by cytometry[J]. Cytometry, 2014, 85(5): 386-399.DOI:10.1002/cyto.a.22452.
[6] 陈婕, 赵茜, 刘贝贝, 等. 交通来源大气污染对慢性阻塞性肺部疾病患者呼吸道氧化应激和炎性反应的影响[J]. 中华预防医学杂志, 2016, 50(5): 411-417. DOI:10.3760/cma.j.issn.0253-9624.2016.05.005.
[7] BREEN A P, MURPHY J A. Reactions of oxyl radicals with DNA[J]. Free Radical Biology and Medicine, 1995, 18: 1033-1077.DOI:10.1016/0891-5849(94)00209-3.
[8] CRUNKHOM S. Neurodegenerative disorders: restoring the balance[J]. Nature Reviews Drug Discovery, 2011, 10(8): 576-585.DOI:10.1038/nrd3521.
[9] SAS K, ROBOTKA H, TOLDI J, et al. Mitochondria, metabolic disturbances, oxidative stress and the kynurenine system, with focus on neurodegenerative disorders[J]. Journal of the Neurological Sciences,2007, 257: 221-239. DOI:10.1016/j.jns.2007.01.033.
[10] KURIEN B T, HENSLEY K, BACHMAVV M, et al. Oxidatively modified autoantigens in autoimmune diseases[J]. Free Radical Biology and Medicine, 2006, 41(4): 549-456. DOI:10.1016/j.freeradbiomed.2006.05.020.
[11] 吴祖芳, 洪松虎, 沈锡权, 等. 乳酸菌高抗氧化活性菌株的筛选及鉴定[J]. 中国食品学报, 2010, 10(1): 73-78. DOI:10.16429/j.1009-7848.2010.01.023.
[12] 刘洋, 郭宇星, 潘道东. 4 种乳酸菌体外抗氧化能力的比较研究[J]. 食品科学, 2012, 33(11): 25-29. DOI:10.13982/j.mfst.1673-9078.2016.12.035.
[13] 陈明, 柯文灿, 张娟, 等. 青藏高原牦牛酸奶中具有抗氧化活性乳酸菌的体内外益生特性[J]. 食品科学, 2017, 38(23): 178-183.DOI:10.7506/spkx1002-6630-201723028.
[14] 侯保朝, 陈丽娥, 陈苏, 等. 功能性乳酸菌的筛选、鉴定及抗氧化性能的初步评价[J]. 食品工业科技, 2015, 36(24): 145-150.DOI:10.13386/j.issn1002-0306.2015.24.023.
[15] CHEN H L, WANG C H, KUO Y W, et al. Antioxidative and hepatoprotective effects of fructo-oligosaccharide in D-galactosetreated Balb/cJ mice[J]. The British Journal of Nutrition, 2011, 105(6):805-809. DOI:10.1017/S000711451000437X.
[16] ITO M, OHISHI K, YOSHIDA Y, et al. Antioxidative effects of lactic acid bacteria on the colonic mucosa of iron-overloaded mice[J].Journal of Agricultural and Food Chemistry, 2003, 51(15): 4456-4460.DOI:10.1021/jf0261957.
[17] ITO M, SAWADA H, OHISHI K, et al. Suppressive effects of bifidobacteria on lipid peroxidation in the colonic mucosa of ironoverloaded mice[J]. Journal of Dairy Science, 2001, 84(7): 1583-1589.DOI:10.3168/jds.S0022-0302(01)74591-2.
[18] WANG Yuzhen, LI Yunxu, XIE Jiming, et al. Protective effects of probiotic Lactobacillus casei Zhang against endotoxin- and D-galactosamine-induced liver injury in rats via anti-oxidative and anti-inf l ammatory capacities[J]. International Immunopharmacology,2013, 15: 30-37. DOI:10.1016/j.intimp.2012.10.026.
[19] HATHOUT A S, MOHAMED S R, EL-NEKEETY A A, et al. Ability of Lactobacillus casei and Lactobacillus reuteri to protect against oxidative stress in rats fed aflatoxins-contaminated diet[J]. Toxicon,2011, 58: 179-186. DOI:10.1016/j.toxicon.2011.05.015.
[20] KAMALADEVI A, GANGULI A, KUMAR M, et al.Lactobacillus casei protects malathion induced oxidative stress and macromolecular changes in Caenorhabditis elegans[J]. Pesticide Biochemistry and Physiology, 2013, 105(3): 213-223. DOI:10.1016/j.pestbp.2013.02.005.
[21] SHARMA R, KAPILA R, KAPASIVA M, et al. Dietary supplementation of milk fermented with probiotic Lactobacillus fermentum enhances systemic immune response and antioxidant capacity in aging mice[J]. Nutrition Research, 2014, 34: 968-981.DOI:10.1111/j.1751-0813.1995.tb03509.
[22] LI S J, HUANG R H, SHAH N P, et al. Antioxidant and antibacterial activities of exopolysaccharides from Bifidobacterium bifidum WBIN03 and Lactobacillus plantarum R315[J]. Journal of Dairy Science, 2014, 97: 7334-7343. DOI:10.3168/jds.2014-7912.
[23] VIIAVAKUMAR M, ILAVENIL S, KIM D H, et al. In-vitro assessment of the probiotic potential of Lactobacillus plantarum KCC-24 isolated from Italian rye-grass (Lolium multif l orum) forage[J].Anaerobe, 2015, 32: 90-97. DOI:10.1016/j.anaerobe.2015.01.003.
[24] WANG Y, ZHOU J Z, XIA X D, et al. Probiotic potential of Lactobacillus paracasei FM-LP-4 isolated from Xinjiang camel milk yoghurt[J]. International Dairy Journal, 2016, 62: 28-34. DOI:10.1016/j.idairyj.2016.07.001.
[25] ZHANG Li, LIU Chunhong, LI Da, et al. Antioxidant activity of an exopolysaccharide isolated from Lactobacillus plantarum C88[J].International Journal of Biological Macromolecules, 2013, 54: 270-275. DOI:10.1016/j.ijbiomac.2012.12.037.
[26] ZHANG D D. Mechanistic studies of the Nrf2-Keap1 signaling pathway[J]. Drug Metabolish Reviews, 2006, 38: 769-789.DOI:10.1080/03602530600971974.
[27] ZHOU Ningna, TANG Yang, KEEP R F, et al. Antioxidative effects of Panax notoginseng saponins in brain cells[J]. Phytomedicine, 2014,21: 1189-1195. DOI:10.1016/j.phymed.2014.05.004.
[28] ARNÉR E S, HOMGREN A. Physiological functions of thioredoxin and thioredoxin reductase[J]. European Journal of Biochemistry, 2000,267(20): 6102-6109. DOI:10.1046/j.1432-1327.2000.01701.x.
[29] NORDBERG J, ARNÉR E S. Reactive oxygen species, antioxidants,and the mammalian thioredoxin system[J]. Free Radical Biology and Medicine, 2001, 31(11): 1287-1312. DOI:10.1016/S0891-5849(01)00724-9.
[30] LI X N, SONG J, ZHANG L, et al. Activation of the AMPK-FOXO3 pathway reduces fatty acid-induced increase in intracellular reactive oxygen species by upregulating thioredoxin[J]. Diabetes, 2009, 58(10):2246-2257. DOI:10.2337/db08-1512.
[31] YAMADA T, IWASAKI Y, NAGATA K, et al. Thioredoxin-1 protects against hyperoxia-induced apoptosis in cells of the alveolar walls[J]. Pulmonary Pharmacology and Therapeutics, 2007, 20(6):650-659. DOI:10.1016/j.pupt.2006.07.00.
[32] TANAKA T, HOSOI F, YAMAGUCHI-IWAI Y, et al. Thioredoxin-2(TRX-2) is an essential gene regulating mitochondria-dependent apoptosis[J]. EMBO Journal, 2002, 21(7): 1695-1703. DOI:10.1093/emboj/21.7.1695.
Mechanism by Which Lactobacillus paracasei FM-M9-1 Repairs D-Galactose-Induced Oxidative Damage in Mice
WANG Ying, XIA Xiudong, DONG Yue, LIU Xiaoli, ZHOU Jianzhong*
(Institute Agro-product Processing, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China)
Abstract:Objective: To analyze the mechanism by which Lactobacillus paracasei FM-M9-1 repairs oxidative damage in mice. Methods: A mouse model of oxidative damage was induced by D-galactose. The model mice were gavaged with strain FM-M9-1 at low and high doses of (5.01 ± 0.05) × 1010and (5.01 ± 0.05) × 108CFU/mL for 60 successive days, separately.Vitamin C was used as a positive control drug. Afterwards, the activity of superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px) and the contents of malondialdehyde (MDA) and protein carbonyl groups in renal and hepatic tissues were measured. We also examined the expression levels of Sod, Gsh-Px, Nrf2 and Trx by real-time fluorescent quantitative polymerase chain reaction (qPCR). Results: The administration of FM-M9-1 significantly enhanced SOD and GSH-Px activities and reduced MDA and protein carbonyl contents. At the transcriptional level, qPCR analysis suggested that the gene expression of Sod, Gsh-Px, Nrf2 and Trx in mice administered with FM-M9-1 increased. Conclusion: Strain FM-M9-1 can alleviate oxidative stress by increasing the activities of antioxidant enzymes, inhibiting lipid and protein oxidation,and up-regulating the expression of antioxidant function-related genes. Overall, strain FM-M9-1 is a promising probiotic candidate for application in functional foods and antioxidant supplements.
Keywords:Lactobacillus paracasei; D-galactose; mice; oxidative damage; repair mechanism
WANG Ying, XIA Xiudong, DONG Yue, et al. Mechanism by which Lactobacillus paracasei FM-M9-1 repairs D-galactose-induced oxidative damage in mice[J]. Food Science, 2018, 39(21): 160-165. (in Chinese with English abstract)DOI:10.7506/spkx1002-6630-201821024. http://www.spkx.net.cn
引文格式:王英, 夏秀东, 董月, 等. 副干酪乳杆菌FM-M9-1对D-半乳糖诱导氧化损伤小鼠的修复机制[J]. 食品科学, 2018, 39(21):160-165. DOI:10.7506/spkx1002-6630-201821024. http://www.spkx.net.cn
文章编号:1002-6630(2018)21-0160-06
文献标志码:A
中图分类号:TS201.3
DOI:10.7506/spkx1002-6630-201821024
*通信作者简介:周剑忠(1965—),男,研究员,博士,研究方向为食品生物技术与生物工程。E-mail:zjzluck@126.com
第一作者简介:王英(1978—),女,副研究员,博士,研究方向为食品生物技术。E-mail:wy116009@126.com
基金项目:国家自然科学基金青年科学基金项目(31501460);南京市科技发展计划项目(201608060)
收稿日期:2017-05-27