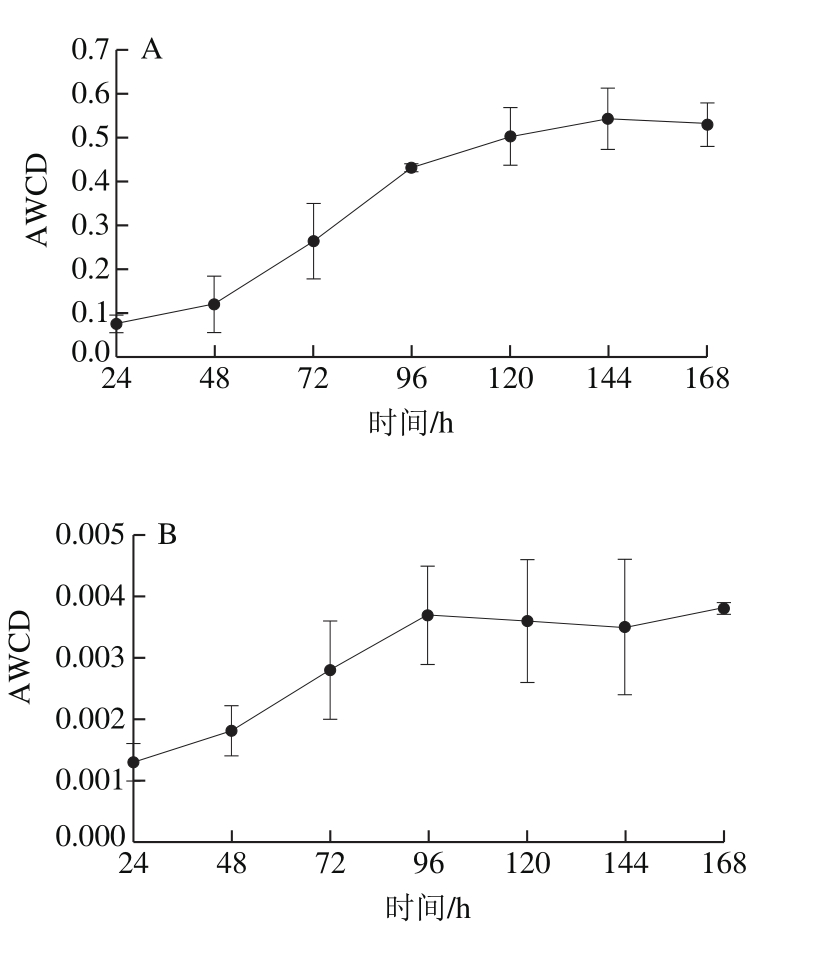
图1 完熟(A)和青熟(B)库车小白杏内生细菌群落AWCD变化
Fig. 1 Changes in AWCD of endophytic bacteria in fully ripe (A) and green ripe (B) apricots
刘晓静1,2,朱 静2,楚 敏2,唐琦勇2,顾美英2,王 博2,朱 璇1,*,张志东1,2,*
(1.新疆农业大学食品科学与药学学院,新疆 乌鲁木齐 830052;2.新疆农业科学院微生物应用研究所,新疆 乌鲁木齐 830091)
摘 要:为分析小白杏采后内生细菌群落的碳源代谢特征和结构组成的变化,实验利用Biolog生态板法及高通量测序法,对其内生细菌群落进行相关分析。结果表明,完熟库车小白杏采后内生细菌群落代谢活性明显高于青熟小白杏,两者主要碳源类型利用存在明显差异。随着采后成熟度的增加,利用氨基酸类、羧酸类、多聚物类内生细菌群落比例明显升高。高通量测序表明,库车小白杏采后内生细菌共包括47 个操作分类单元,涉及15 个目,33 个属,两者内生细菌群落结构差异明显;黄杆菌目(Flavobacteriales)、拟杆菌目(Bacteroidales)、梭菌目(Clostridiales)占青熟小白杏内生细菌群落的83%;完熟小白杏内生细菌群落的主要组成为红螺菌目(Rhodospirillales)、肠杆菌目(Enterobacteriales)、梭菌目,总计占总体的86%。且随着成熟度的增加,黄杆菌目、拟杆菌目所占比例逐渐下降,而红螺菌目、肠杆菌目所占比例逐渐上升,但梭菌目未见明显差异。同时,研究发现小白杏内生细菌中存在水果软腐病和人体肠胃条件致病等相关菌群,为进一步阐明库车小白杏采后腐败变质内在机理,开发相关的贮存、保鲜方法及其科学合理食用等提供一定研究基础。
关键词:库车小白杏;内生细菌;代谢特征;群落变化
库车小白杏,蔷薇科,李亚科,因盛产于新疆阿克苏地区库车县而得名[1]。库车小白杏汁多味鲜、风味浓郁,果肉富含丰富的胡萝卜素和VC,且含糖高,是优良的鲜食水果[2-3]。同时,其果肉还可以加工为果醋,其仁可加工成高附加值的油脂,其种子被用来治疗皮肤病以及咳嗽、哮喘、便秘,已成为本地重要经济林果之一[4-8]。
由于库车小白杏鲜果成熟期集中,贮存期短,易腐败变质,一般鲜果销售期仅为3~5 d,严重制约了库车小白杏的生产和销售。研究表明,库车小白杏是典型的呼吸跃变型果实,采后果肉和果皮易褐变,果实易软化、腐烂和霉变。即使在0 ℃冷藏,库车小白杏也会发生后熟,冷藏18~27 d果实转为完熟期,之后进入衰老期,果实出现软腐,进而产生霉变[9]。目前,有关库车小白杏的研究主要集中于其采后生理变化和保鲜、贮存等方面[10-12],而有关库车小白杏内生菌的研究尚鲜见报道。作为多种途径参与植物代谢、发育的内生菌,其与宿主植物在长期的系统演化过程中形成了协同进化关系[13-14]。目前,国内已对枸杞、枣、柑橘、葡萄、芒果、哈密瓜等果实内生菌进行了研究,发现其中存在丰富的内生菌,呈现出一定的寄主偏好性,且蕴藏着宝贵的天然产物产生菌和病原拮抗菌。胡美姣等[15]对采后芒果果实的研究发现,大多病原菌具有潜伏侵染的特性,如可可球二孢菌在果实未成熟时就会浸染,导致果实在贮运及销售过程中发生快速且严重的腐烂。
本研究以库车小白杏为实验材料,通过Biolog生态板法和高通量测序法,对采后不同时期其内生细菌群落碳源代谢特征及结构组成变化进行研究,以期从内生细菌的角度阐明库车小白杏易腐变质的机制,为进一步的研究提供理论依据。
青熟小白杏采自库车县乌恰镇小白杏果园中,其表皮青涩,果实硬度较高,无机械伤痕,无病变;完熟小白杏为采摘的青熟小白杏于室温、避光条件下放置7 d,至其表皮为橘色,果实较软,无机械伤痕,无病变。
生态板、自动微生物鉴定系统 美国Biolog公司。
1.3.1 样品的预处理
分别取青熟和完熟小白杏各20 颗,称质量后经自来水流水冲洗10 min,无菌纸吸干水分,75%无水乙醇处理4 min,经无菌水洗涤后,用3% H2O2浸泡3 min,再用无菌水冲洗3~4 次,最后一遍无菌水洗涤液经涂板检测无菌后,于无菌条件下进行库车小白杏的去核、破碎、匀浆等操作。
1.3.2 内生菌群落碳源代谢利用分析
采用Biolog生态板法进行。库车小白杏经表面完全消毒后,无菌条件下称取20 g果肉,以无菌水-果肉比9∶1的比例再次进行匀浆,静置30 min后,每孔取上清液150 μL接种至生态板,每个处理3 个重复。于28 ℃恒温培养,每隔24 h分别读数,连续培养7 d。
以微平板孔平均颜色变化率(average well color development,AWCD)(吸光度表示)进行库车小白杏采后内生细菌群落代谢活性的分析,采用DPS v9.50版软件进行内生菌群落碳源利用率及多样性分析[16]。
1.3.3 内生菌总DNA的提取
经表面消毒的库车小白杏,采用十六烷基三甲基溴化铵(cetyltrimethylammonium bromide,CTAB)法对样本的基因组DNA进行大量提取[17],经凝胶电泳检测后,DNA浓度及纯度合格后作为聚合酶链式反应(polymerase chain reaction,PCR)扩增模板使用。
1.3.4 16S rDNA序列V4可变区的PCR扩增
以前述内生菌DNA为模板,采用引物为515F和806R,扩增16S rDNA序列的V4可变区,PCR条件:94 ℃ 5 min;94 ℃ 30 s,60 ℃ 30 s,72 ℃ 30 s,30 个循环;72 ℃ 10 min。扩增PCR产物经电泳检测切胶回收,并检测浓度调至一致后,样品送至北京诺禾致源生物信息科技有限公司进行高通量测序。
1.3.5 数据预处理
所得的测序数据经去除冗余和无效序列后,对有效序列Tags聚类,以同源性大于97%聚为一个操作分类单元(operational taxonomic unit,OTU),作为一个假定的分类单元。选取代表性序列,通过RDP classifier软件进行物种分类,确定OTU之间的分类地位。
1.3.6 Alpha多样性分析
Alpha多样性分析采用DPS v9.50版软件,分别采用覆盖率、物种丰富度、物种多样性等方面信息评价。其中,覆盖率用来反映测序深度;OTU用来分析群落种群丰富度,香农指数和辛普森指数用来估算微生物群落的多样性[18-19]。
2.1.1 AWCD分析
样品经7 d连续培养和测定,数据经分析并求得平均吸光度后作图,由图1可以看出,青熟和完熟小白杏各处理的平均吸光度有明显区别,完熟小白杏处理在培养48~120 h内,表现出快速增长的趋势,其后趋于缓慢,其最大吸光度为0.543;而青熟小白杏处理在24~96 h间略有增加后,便不再增加,其最大吸光度为0.003 8。结果表明,完熟库车小白杏内生菌群落代谢活性远高于青熟小白杏,约为青熟小白杏的143 倍。
图1 完熟(A)和青熟(B)库车小白杏内生细菌群落AWCD变化
Fig. 1 Changes in AWCD of endophytic bacteria in fully ripe (A) and green ripe (B) apricots
2.1.2 内生菌群落碳源利用分析
根据AWCD变化情况,研究选择培养96 h时内生菌群落对31 种底物利用情况,并将其归类为6 类碳源,以分析各样品的碳源利用情况。由图2可以看出,库车小白杏青熟和完熟两个阶段内生细菌群落对于6 大碳源种类的利用存在明显差异。在青熟小白杏中,碳水化合物类和胺类为其内生菌群落主要利用碳源类型,分别占总体的60%和14%;而在完熟小白杏中,碳水化合物类和羧酸为群落主要利用类型,分别占总体的30%和22%。比较分析发现,随着成熟度的增加,小白杏内生菌群落利用碳水化合物类、酚酸类和胺类内生菌群落比例明显降低,而利用氨基酸类、羧酸类、多聚物类内生细菌群落比例升高。
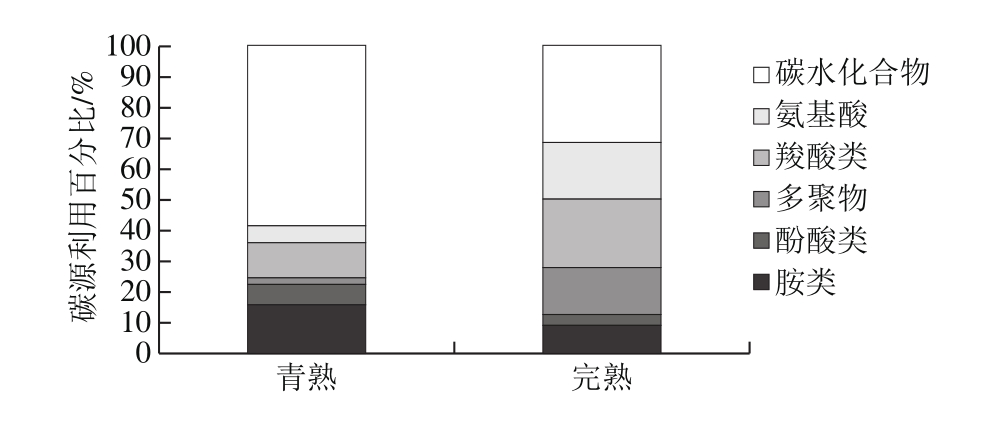
图2 青熟和完熟库车小白杏内生细菌群落对不同类型碳源利用
Fig. 2 Carbon source utilization profiles of endophytic bacterial communities in fully ripe and green ripe apricots

图3 青熟(A)和完熟(B)小白杏内生细菌群落主要利用的8 种碳源
Fig. 3 Utilization rates of 8 major carbon sources by endophytic bacterial communities in green ripe (A) and fully ripe (B) apricots
进一步对31种碳源的具体利用情况分析表明,完熟小白杏内生菌群落利用碳源数量明显高于青熟小白杏,其中完熟小白杏所利用的11 种碳源未在青熟小白杏处理中发现。各碳源利用所占比例分析表明,D-半乳糖酸-γ-内脂、腐胺、D-木糖、α-D-乳糖、D-甘露醇、γ-羟丁酸、2-羟基苯甲酸、4-羟基苯甲酸为青熟小白杏内生菌群落主要利用碳源(图3A);而丙酮酸甲酯、α-丁酮酸、吐温40、吐温80、苯乙胺、L-精氨酸、D-纤维二糖、衣康酸为完熟小白杏主要利用碳源(图3B)。综上证明,青熟和完熟库车小白杏采后内生菌群落结构发生了明显变化。
2.2.1 高通量测序数据预处理
通过各样品中内生细菌的16S rDNA V4区测序,2组样品总计测得原始序列106 317 条,过滤掉低质量序列后,总数为104 286 条。所得序列经聚类比对,以相似度低于97%以聚类为1 个OTU,并去除植物体内的线粒体、叶绿体相关序列后,共获得47 个OTU,各样品测序信息见表1。
表1 青熟和完熟库车小白杏微生物菌株测序数据统计
Table 1 Statistics of sequencing data for microbial strains in fully ripe and green ripe apricots

2.2.2 样品间Alpha多样性分析
通过覆盖率、物种丰富度、物种多样性等方面信息对各样品间Alpha多样性进行了描述,见表2。结果表明,各样品所得序列覆盖了全部内生菌的99%,所得数据能够准确地反映出库车小白杏采后内生菌群落的组成;OTU分析表明,库车小白杏采后内生菌群落的组成单元变化不大;但完熟小白杏内生菌群落香农指数及辛普森指数较青熟小白杏略有降低,说明群落的均一性发生变化,完熟小白杏内生菌群落的有一定的分化。
表2 不同成熟度库车小白杏微生物多样性统计
Table 2 Bacterial diversity in fully ripe and green ripe apricots

2.2.3 内生菌群落分布特征分析
利用RDP classifie对各样品中OTU依次进行门、纲、目、科、属分类信息分析,进一步挖掘样品的群落组成。结果表明,库车小白杏采后内生细菌群落共涉及8 个门,12 个纲,15 个目,23 个科,33 个属。
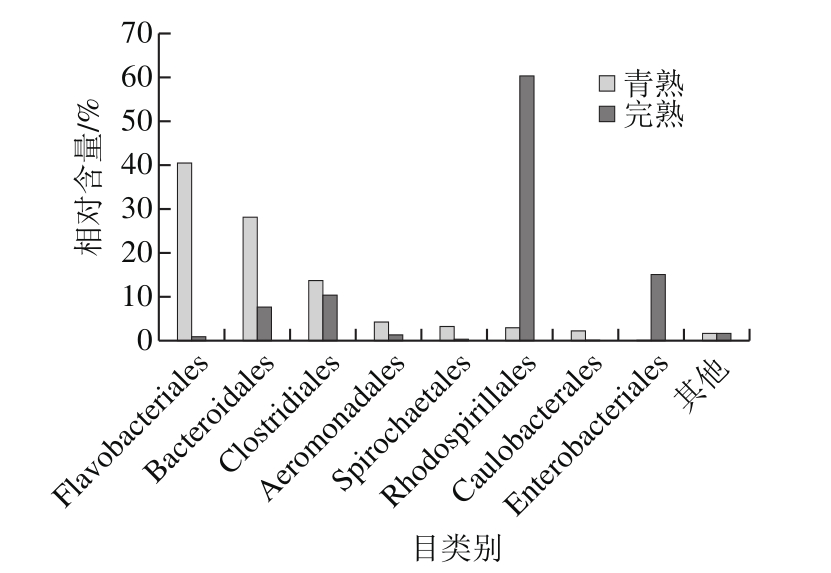
图4 目水平上主要的库车小白杏内生细菌群落组成
Fig. 4 Major bacterial orders in fully ripe and green ripe apricots apricot
通过对样品中主要目的分析发现,青熟和完熟小白杏采后内生细菌群落的分布比例存在明显差异。如图4所示,青熟小白杏内生细菌群落的主要组成为黄杆菌目(Flavobacteriales)、拟杆菌目(Bacteroidales),分别占总体的41%、28%,其在完熟小白杏中比例明显下降;红螺菌目(Rhodospirillales)、肠杆菌目(Enterobacteriales)为完熟小白杏内生细菌群落的主要组成,分别占总体的60%、15%,其较青熟小白杏内生细菌群落中所占比例上升明显;梭菌目在采后小白杏内生菌中比例变化不大,仅由青熟中的10%提高至完熟的14%。而其他菌如如气单胞菌目(Aeromonadales)、螺旋体目(Spirochaetales)等非优势菌群均随着小白杏的成熟度的增加有所下降。
表3 属水平上主要的库车小白杏内生细菌群落组成
Table 3 Major bacterial genera in fully ripe and green ripe apricots apricota

进一步分析发现,库车小白杏采后内生细菌群落共涉及33 个属,其中相对含量大于0.01%的菌属,青熟小白杏涉及14 个,完熟小白杏涉及12 个,表明随着小白杏成熟度的增加,其内生菌群的均匀度下降,这一结果与Biolog生态板法分析结果一致。通过对样品中主要菌属分析发现(表3),青熟小白杏的主要内生菌群由蟑螂杆状体科未分类属、普氏菌属和瘤胃球菌科NK4A214类群微生物组成,分别占总体的50.60%、9.64%和6.42%,约占总体的67%,但随着小白杏成熟度的增加,其群落所占比例明显下降,仅占完熟小白杏内生细菌的1.39%、3.65%、2.09%;葡糖杆菌属为完熟小白杏的绝对优势菌群,占总体的74.09%,其次为泛菌属、克里斯滕森菌科R-7类群、普氏菌属,分别占总体的4.17%、3.83%、3.65%。随着小白杏成熟度的增加,葡糖杆菌属所占比例增加了近17.5 倍,泛菌属增加了约416 倍,而蟑螂杆状体科未分类属降低了约97.25%,普氏菌属和瘤胃球菌科NK4A214菌群降低了约67.45%。
由上述可见,采后完熟库车小白杏的内生细菌群落组成明显有别于青熟小白杏,且随着采后成熟度的增加,其菌群组成结构存在着明显的更替。
植物内生菌种类繁多,可广泛分布于不同植物的不同部位,能产生多种活性代谢产物,如具有抗肿瘤、抗菌、抗病毒、杀虫、免疫抑制、抗氧化和降糖等活性功能[20-24],已成为重要的活性功能产物的来源。同时,内生菌也可通过多种途径参与植物的代谢、发育和成熟,是研究微生物与植物协同进化的重要研究材料。此前,国内外对植物果实内生菌的研究较少,特别是对新疆特色水果内生菌的研究尚鲜见报道。本研究利用Biolog生态板法及高通量测序法,对库车小白杏采后内生细菌群落变化进行分析。研究表明,库车小白杏采后内生细菌群落代谢活性发生了明显变化,完熟库车小白杏内生菌群落代谢活性远高于青熟小白杏,约为青熟小白杏的143 倍。进一步分析表明,随着库车小白杏成熟度的增加,利用碳水化合物类、酚酸类和胺类内生菌群落比例明显降低,而利用氨基酸类、羧酸类、多聚物类内生细菌群落比例升高。上述结果,可能与库车小白杏属于呼吸跃变型果实有关。
同时,对其采后内生细菌群落组成变化分析表明,小白杏内生细菌共包括47 个OTU,涉及15 个目,33 个属。在青熟和完熟小白杏中,其两者内生细菌群落结构差异明显。在目一级的分类水平上,黄杆菌目、拟杆菌目和梭菌目菌株是青熟小白杏内生细菌群落主要组成,占总菌群种类的83%;而完熟小白杏内生细菌群落主要由红螺菌目、肠杆菌目和梭菌目菌群组成,占总体的86%。随着小白杏采后成熟度的增加,黄杆菌目、拟杆菌目菌群所占比例逐渐下降,而红螺菌目、肠杆菌目菌群所占比例逐渐上升,梭菌目菌群变化不大。在属一级的分类水平上分析表明,随着小白杏成熟度的增加,其内生菌群的均匀度下降,这一结果与Biolog生态板法分析结果相比分别提高了17.5 倍和416 倍,而蟑螂杆状体科未分类属所占比例下降最为明显,差异约达97.25%;另外,普氏菌属和瘤胃球菌科NK4A214菌群也降低了约67.45%。由上述可见,采后完熟库车小白杏的内生细菌群落组成明显有别于青熟小白杏,且随着采后成熟度的增加,其菌群组成结构存在着明显的更替。
现有研究表明,植物内生菌结构组成易受寄主生长发育而变化。郑惠文等[24]对新疆杏果实发育过程中可溶性糖和有机酸变化的研究发现,杏果实中更易被细菌所利用的葡萄糖明显下降,而蔗糖的含量明显增加,这可能与完熟小白杏内生细菌中利用碳水化合物菌群所占比例下降的有关。而随着杏果实的成熟度的增加,果实中总酸下降,有机酸由苹果酸积累型向苹果酸、奎宁酸和柠檬酸共积累的模式转变,造成果实中pH值上升,有机酸种类的增加[25-26],可能刺激了利用羧酸类细菌的增长。同时,随着成熟度的增加,果肉中果胶、可溶性淀粉等易分解多聚物类化合物的增加,以及营养成分中氨基酸的增加及溶出,均可能导致多聚物类、氨基酸类内生细菌群落比例升高。而胡美姣[14]对采后芒果果实的研究发现,大多病原菌具有潜伏侵染的特性,会导致果实在采后贮运及销售过程中发生快速腐烂。本研究发现,小白杏内生细菌中葡糖杆菌属、泛菌属等相关植物软腐病原菌在完熟期含量明显上升,可能与库车小白杏采后的软化腐败有关[26-30],相关验证工作正在开展。
另外,中国自古以来就有“杏伤人”的说法,即过多食用杏子对身体不利。通过对菌群分析发现,小白杏果肉内生细菌中存在着多种与肠胃代谢相关菌群。如拟杆菌、克氏杆菌、理研菌科、瘤胃菌科、普雷沃氏菌属等菌群,虽广泛存在健康人群的口腔或肠道中,但当机体免疫功能紊乱或菌群失调时,如拟杆菌、普雷沃氏菌等能导致内源性感染;而气单胞菌属和密螺旋体属的菌群,可引起人体腹泻等多种感染,在国外作为腹泻病原菌进行检测[31-37]。
另一方面,杏有降气止咳平喘、润肠通便的功效。本研究表明,库车小白杏内生菌中也存在多种相关有益菌群,如葡糖杆菌,可促进VC的合成,是合成降血糖药物-米格列醇的关键中间体[34-35];库车小白杏内生菌中存在着多种克里斯滕森菌、拟杆菌、瘤胃杆菌及理研菌等,其均与人体肥胖呈负相关[26,38]。有研究[39-40]发现,克里斯滕森菌科菌群可有效改变人和老鼠等宿主代谢的方式,从而降低动物个体的体重指数。刘璨[34]研究发现拟杆菌菌群丰度与肥胖指数呈反比,其在健康人体中的含量明显高于肥胖患者;通过对实验小鼠进行饲喂拟杆菌菌液,可对肥胖大鼠有一定的减肥作用。相关研究[41-43]表明瘤胃菌科与脂肪因子水平具有明显的相关性,在能量代谢中占主导地位。由此可见,杏的多种功能和功效可能与其内生细菌群落有关,相关论点仍需更多的杏品种内生菌研究的验证。
上述研究结果不仅为库车小白杏采后腐败变质内在机理提供了基础,为其贮存、保鲜提供了科学借鉴,也为阐释小白杏营养学和医学中功能的研究提供了理论依据。
参考文献:
[1] 赵多勇, 康露, 王成, 等. 新疆杏子生产现状及存在的问题[J]. 农业工程, 2016, 6(5): 53-56.
[2] 赵翠, 田英姿, 英犁, 等. 新疆杏和华北杏果实品质的比较分析[J]. 现代食品科技, 2014, 30(7): 286-291. DOI:10.13982/j.mfst.1673-9078.2014.07.052.
[3] 张君萍, 高疆生, 李疆, 等. 新疆杏与华北杏果实主要营养成分比较分析[J]. 新疆农业科学, 2006, 43(2): 140-144.
[4] LEE H H, AHN J H, KWON A R, et al. Chemical composition and antimicrobial activity of the essential oil of apricot seed[J]. Phytotherapy Research, 2015, 28(12): 1867-1872. DOI:10.1002/ptr.5219.
[5] 武璇. 新疆库车小白杏杏仁油调节大鼠血脂作用及机制的实验研究[D]. 乌鲁木齐: 新疆医科大学, 2012: 1-7.
[6] 陈新军, 唐凤仙, 单春会, 等. 库车小白杏果醋饮料工艺研究[J].食品工业, 2016, 37(8): 171-175.
[7] 程卫东, 吕国华, 李琳, 等. 新疆小白杏资源的综合利用及其产业化[J].农业工程学报, 2006, 22(9): 241-244.
[8] 温昕晔. 一氧化氮对小白杏冷藏特性及有机酸代谢相关基因表达的研究[D]. 乌鲁木齐: 新疆农业大学, 2015: 1-3.
[9] 张辉, 李洁, 温昕晔, 等. 冷藏期间小白杏果实后熟过程中相关抗逆特性的表现[J]. 保鲜与加工, 2013, 13(4): 15-18.
[10] 迪力夏提·卡迪尔. 冷激处理对小白杏贮藏品质及生理效应的影响[D].乌鲁木齐: 新疆农业大学, 2013: 1-7.
[11] 王聘. 新疆小白杏果实采后贮藏保鲜的研究[D]. 南京: 南京农业大学, 2012: 1-21.
[12] 王莉莉. 入侵植物南美蟛蜞菊内生细菌多样性及其生态学潜在功能研究[D]. 镇江: 江苏大学, 2013: 1-11.
[13] 陈龙, 梁子宁, 朱华. 植物内生菌研究进展[J]. 生物技术通报, 2015,31(8): 30-34. DOI:10.13560/j.cnki.biotech.bull.1985.2015.08.005.
[14] 胡美姣. 芒果果实潜伏侵染、Botryodiplodia theobromae致腐机理及蒂腐病防治技术基础研究[D]. 海口: 海南大学, 2013: 1-18.
[15] 姚槐应, 黄昌勇. 土壤微生物生态学及其实验技术[M]. 北京: 科学出版社, 2006.
[16] 黄雅丽, 马嫚, 张韧, 等. DNA提取方法对桑树内生菌多样性研究的影响[J]. 广州中医药大学学报, 2015, 32(4): 729-734. DOI:10.13359/j.cnki.gzxbtcm.2015.04.033.
[17] 张志东, 顾美英, 王玮, 等. 基于高通量测序的辐射污染区细菌群落特征分析[J]. 微生物学通报, 2016, 43(6): 1218-1226. DOI:10.13344/j.microbiol.china.150371.
[18] 顾美英, 徐万里, 茆军, 等. 新疆绿洲农田不同连作年限棉花根际土壤微生物群落多样性[J]. 生态学报, 2012, 32(10): 3031-3040.
[19] 黄敬瑜, 张楚军, 姚瑜龙, 等. 植物内生菌生物抗菌活性物质研究进展[J]. 生物工程学报, 2017, 33(2): 178-186. DOI:10.13345/j.cjb.160277.
[20] DONMEZ M F, KARLIDAG H, ESITKEN A. Identification of resistance to bacterial canker (Pseudomonas syringae pv. syringae)disease on apricot genotypes grown in Turkey[J]. European Journal of Plant Pathology, 2010, 126(2): 241-247. DOI:10.1007/s10658-009-9536-x.
[21] YAO T, ZHOU Y, CHANGYONG A Z. Research development of the differentiation and control of citrus bacterial canker disease[J]. Science &Technology of Food Industry, 2011, 32(12): 139-141.
[22] MORYAN G. Pathogenic organisms as causes of apricot decline[J]. Acta Horticulturae, 1995, 384: 521-532. DOI:10.17660/ActaHortic.1995.384.82.
[23] CRUZ A F, SOARES W R D O, BLUM L E B. Impact of the arbuscular mycorrhizal fungi and bacteria on biocontrol of white root rot in fruit seedlings[J]. Plant Physiology & Pathology, 2014, 2: 1-5.DOI:10.4172/2329-955X.1000114.
[24] 郑惠文, 张秋云, 李文慧, 等. 新疆杏果实发育过程中可溶性糖和有机酸的变化[J]. 中国农业科学, 2016, 49(20): 3981-3992.
[25] 陈美霞, 赵从凯, 陈学森, 等. 杏果实发育过程中糖积累与蔗糖代谢相关酶的关系[J]. 果树学报, 2009, 26(3): 320-324. DOI:10.13925/j.cnki.gsxb.2009.03.012.
[26] 冯静, 施庆珊, 欧阳友生, 等. 葡糖杆菌属分类及其主要应用的研究进展[J]. 微生物学杂志, 2010, 30(2): 86-90.
[27] 曹慧英, 李洪杰, 朱振东, 等. 玉米细菌干茎腐病菌成团泛菌的种子传播[J]. 植物保护学报, 2011, 38(1): 31-36. DOI:10.13802/j.cnki.zwbhxb.2011.01.004.
[28] 成文玉, 金红星, 胡炎华, 等. 明串珠菌筛选与分类的研究进展[J].中国酿造, 2010, 29(3): 7-9.
[29] 王晓燕, 桑卫国, 章宗铭. 腐烂紫菜中琼胶酶高产菌株的筛选及鉴定[J]. 食品科技, 2009, 34(7): 2-5. DOI:10.13684/j.cnki.spkj.2009.07.038.
[30] 韩少锋. 罗非鱼腐败过程菌群结构分析及腐败菌的分离、鉴定与调控[D]. 北京: 中国农业科学院, 2010: 43-46.
[31] 智发朝, 白杨, 王晔, 等. 一种脆弱拟杆菌在预防和/或治疗炎症性肠病中的应用: CN105434476A[P]. 2016-03-30.
[32] SILVA L C A D, LEAL-BALBINO T C, MELO B S T D, et al.Genetic diversity and virulence potential of clinical and environmental Aeromonas spp. isolates from a diarrhea outbreak[J]. Bmc Microbiology,2017, 17(1): 179-188. DOI:10.1186/s12866-017-1089-0.
[33] 谢婧雯, 王烨, 朱明, 等. 多形拟杆菌对糖尿病模型小鼠的影响[J].中国微生态学杂志, 2013, 25(8): 869-873. DOI:10.13381/j.cnki.cjm.2013.08.035.
[34] 刘璨. 肥胖易感和肥胖抵抗小鼠的血清代谢组学与肠道菌群的研究[D]. 南昌: 南昌大学, 2016: 27-42.
[35] JOHNSON E L, HEAVER S L, WALTERS W A, et al. Microbiome and metabolic disease: revisiting the bacterial phylum Bacteroidetes[J].Journal of Molecular Medicine, 2017, 95(1): 1-8. DOI:10.1007/s00109-016-1492-2.
[36] 孙锡宝. 猪密螺旋体属痢疾的流行、诊断和防治措施[J]. 兽医导刊,2016(20): 111.
[37] HUWS S A, KIM E J, LEE M R F, et al. As yet uncultured bacteria phylogenetically classified as Prevotella, Lachnospiraceae incertae sedis and unclassified Bacteroidales, Clostridiales and Ruminococcaceae may play a predominant role in ruminal biohydrogenation[J]. Environmental Microbiology, 2011, 13(6): 1500-1512. DOI:10.1111/j.1462-2920.2011.02452.x.
[38] GOMEZARANGO L F, BARRETT H L, MCINTYRE H D, et al.Connections between the gut microbiome and metabolic hormones in early pregnancy in overweight and obese women[J]. Diabetes, 2016,65(8): 2214. DOI:10.2337/db16-0278.
[39] LEY R E, GOODRICH J, WATERS J. Modulation of fat storage in a subject by altering population levels of Christensenellaceae in the GI tract: US2017042948[P]. 2017-02-16.
[40] GOODRICH J K, WATERS J L, POOLE A C, et al. Human genetics shape the gut microbiome[J]. Cell, 2014, 159(4): 789-799.DOI:10.1016/j.cell.2014.09.053.
[41] FERGUSON R, COULON F, VILLA R. Understanding microbial ecology can help improve biogas production in AD[J]. Science of the Total Environment, 2018, 642: 754-763. DOI:10.1016/j.scitotenv.2018.06.007.
[42] ZHAO J B, LIU P, HUANG C F, et al. Effect of wheat bran on apparent total tract digestibility, growth performance, fecal microbiota and their metabolites in growing pigs[J]. Animal Feed Science &Technology, 2018, 239: 14-26. DOI:10.1016/j.anifeedsci.2018.02.013.
[43] SHANG Q, SHAN X, CAI C, et al. Dietary fucoidan modulates the gut microbiota in mice by increasing the abundance of Lactobacillus and Ruminococcaceae[J]. Food & Function, 2016, 7(7): 3224-3232.DOI:10.1039/C6FO00309E.
Changes in Carbon Metabolism Characteristics and Community Composition of Endophytic Bacteria in Postharvest Kuqa-Grown Apricot
LIU Xiaojing1,2, ZHU Jing2, CHU Min2, TANG Qiyong2, GU Meiying2,WANG Bo2, ZHU Xuan1,*, ZHANG Zhidong1,2,*
(1. College of Food Sciences and Pharmacy, Xinjiang Agricultural University, Ürümqi 830052, China;2. Institute of Microbiology, Xinjiang Academy of Agricultural Sciences, Ürümqi 830091, China)
Abstract:This study aimed to analyze the succession of endophytic bacterial communities in postharvest Kuqa-grown apricot. The characteristics of carbon metabolism and the bacterial community composition were investigated by Biolog Eco method and high-throughput sequencing. The results showed that the metabolic activity of endophytic bacteria in Kuqagrown apricot was obviously higher at the fully ripe stage than at the green ripe stage, and the major carbon sources utilized by the endophytic bacteria were significantly different between the two stages. The percentages of endophytic bacterial communities utilizing amino acids, carboxylic acids and polymeric substances were increased markedly. High-throughput sequencing results showed that 47 operational taxonomic units (OTUs) including 33 genera in 15 orders were obtained. The community structure of endophytic bacteria was different significantly in fully ripe and green ripe apricots. Flavobacteriales,Bacteroidales Clostridiales together accounted for up to 83% of the endophytic bacterial community. Meanwhile,Rhodospirillales, Enterobacteriales and Clostridiales were predominant in fully ripe apricot, accounting for up to 86% the total bacterial community. The percentages of Flavobacterium and Bacteroides were decreased whereas the percentages of Rhodospirillales and Enterobacteriales were increased; however, the percentage of Clostridiales did not significantly change with ripening of apricots. Furthermore, some pathogens that caused fruit soft rot and gastrointestinal disorders in humans were observed. This study can provide a scientific basis to further understand the spoilage mechanism of postharvest Kuchegrown apricot and develop storage and preservation technologies and for proper consumption.
Keywords:Kuqa-grown apricot; endophytic bacteria; carbon metabolism characteristics; bacterial community succession
LIU Xiaojing, ZHU Jing, CHU Min, et al. Changes in carbon metabolism characteristics and community composition of endophytic bacteria in postharvest Kuqa-grown apricot[J]. Food Science, 2018, 39(22): 141-146. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201822022. http://www.spkx.net.cn
引文格式:刘晓静, 朱静, 楚敏, 等. 库车小白杏采后内生细菌群落代谢特征及结构组成的变化[J]. 食品科学, 2018, 39(22):141-146. DOI:10.7506/spkx1002-6630-201822022. http://www.spkx.net.cn
文章编号:1002-6630(2018)22-0141-06
文献标志码:A
中图分类号:Q939.1
DOI:10.7506/spkx1002-6630-201822022
张志东(1977—),男,研究员,硕士,研究方向为特殊环境微生物学。E-mail:zhangzheedong@sohu.com
朱璇(1971—),女,教授,博士,研究方向为果蔬采后生理与贮藏保鲜。E-mail:zx9927@126.com
*通信作者简介:
第一作者简介:刘晓静(1994—),女,硕士研究生,研究方向为果蔬贮藏及物流工程。E-mail:liuxiaojing1122@qq.com
基金项目:国家自然科学基金面上项目(31060414);新疆农业科学院农业科技创新平台能力提升建设专项(XJNKYPT-2017-002)
收稿日期:2017-10-11