原发性草酸钙肾结石的形成通常与高钙尿症、高草酸尿症和低枸橼酸尿症有关[1],其中高草酸尿症是草酸钙肾结石形成的主要因素[2],其主要由草酸盐在体内的产生、吸收与分泌失衡导致。机体内的草酸盐主要有两方面的来源,即内源和外源。内源是通过哺乳动物肝脏代谢产生;外源则是通过胃肠道对食源草酸盐的吸收获取,而草酸盐是大多数食源性植物产生的次生代谢产物。哺乳动物体内缺乏分解利用草酸盐的酶类,草酸盐主要是通过尿液排出体外[3]。健康机体每天通过尿分泌的草酸为10~40 mg,如果超过40~45 mg,临床诊断即为高草酸尿症[4]。随着组学技术的发展,肠道微生物的功能逐渐被揭示,例如肠道微生物菌群失衡可导致肾脏疾病,肾脏疾病患者肠道微生物组成也在发生改变,二者是相互的[5]。肠道菌群是影响肠道草酸盐分解的一个重要因素,其中食草酸杆菌为研究最广泛、最深入的草酸盐降解菌[6],并已有相应商业化产品上市[7]。食草酸杆菌在肠道内的定植与尿草酸浓度的降低存在直接相关性[8-11],然而,这种细菌作为益生菌应用时往往受到在肠道定植不稳定和对抗生素敏感的限制[7,12]。近年来一系列体外实验、动物模型实验、人体临床实验均发现,乳酸菌有一定的潜在降解草酸盐的能力,可降解草酸盐的乳酸菌主要包括乳杆菌属、双歧杆菌属和肠球菌属菌株[13-19]。其中,乳杆菌属和双歧杆菌属为人体肠道内固有的有益菌群,通常认为其应用于人体是安全的,因此被广泛作为益生菌应用。本文主要对可降解草酸盐的乳酸菌种类、乳酸菌降解草酸盐的能力以及乳酸菌降解草酸盐的机理进行综述,为降解草酸盐乳酸菌的筛选及其作用机理研究提供指导,也为预防和治疗高草酸尿症益生菌产品的开发提供理论参考。
1 具有降解草酸盐功能的乳酸菌
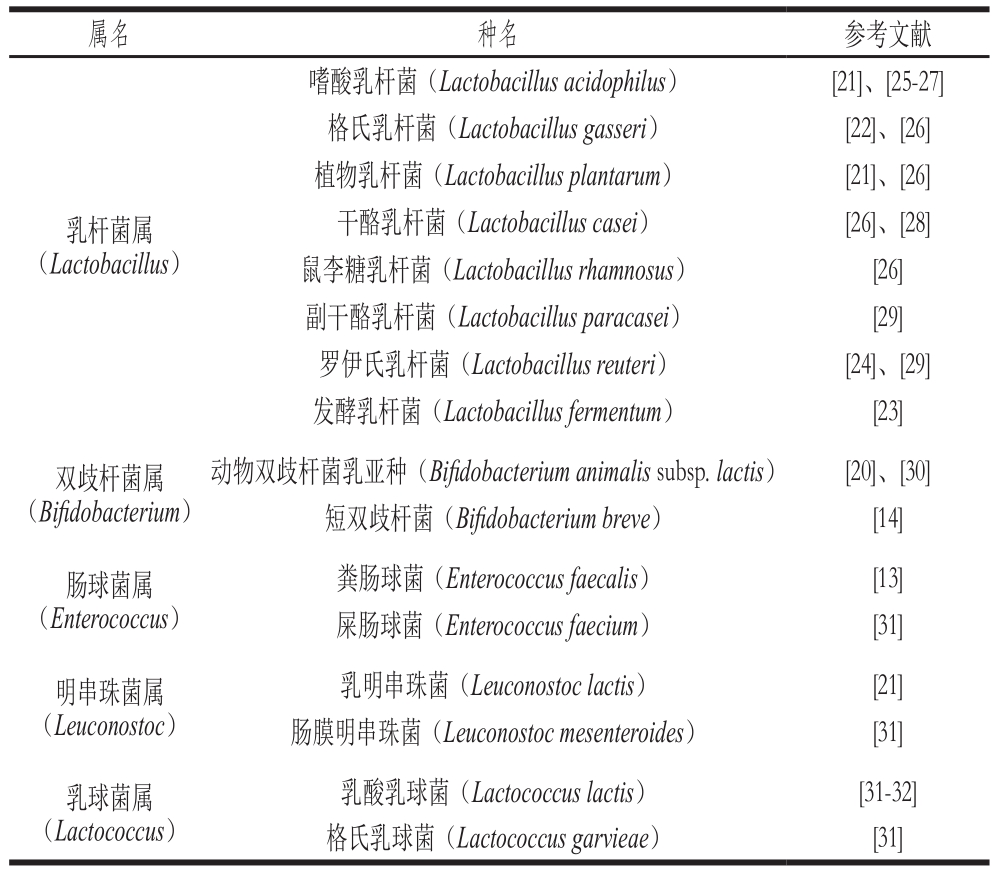
乳酸菌在自然界中分布广泛,传统发酵食品、动物和人的口腔、肠道及阴道中均栖居着大量乳酸菌。国内外研究人员已从自然环境中分离筛选到大量可降解草酸盐的乳酸菌菌株,这些乳酸菌分离株主要来源于传统发酵食品(如酸奶)、人体粪便以及动物粪便样品等[20-24]。目前,已表现出可降解草酸盐活性的乳酸菌主要包括乳杆菌属、双歧杆菌属、肠球菌属、明串珠菌属以及乳球菌属等,其中已明确具有降解草酸盐能力的乳杆菌属中主要有嗜酸乳杆菌[21,25-27]、格氏乳杆菌[26]、植物乳杆菌[21,26]、干酪乳杆菌[26,28]、鼠李糖乳杆菌[26]、副干酪乳杆菌[29]、罗伊氏乳杆菌[24,29]、发酵乳杆菌[23]等,双歧杆菌属主要有动物双歧杆菌乳亚种[20,30]、短双歧杆菌[14]、肠球菌属中的粪肠球菌[13]和屎肠球菌[31]、明串珠菌属中的乳明串珠菌[21]和肠膜明串珠菌[31],以及乳球菌属中的乳酸乳球菌[31-32]和格氏乳球菌[31],具体见表1。除此之外,嗜热链球菌、德氏乳杆菌保加利亚亚种、短乳杆菌、婴儿双歧杆菌、长双歧杆菌的个别菌株可能也具有降解草酸盐的活性,主要是因为这些菌一般与其他乳酸菌联合制成益生菌制剂应用,如Oxadrop®和VSL#3®产品[33-34]。另外,魏斯氏菌属内的一些菌株也展现出了一定的草酸盐降解活性[23]。乳酸菌降解草酸盐的活性具有菌种特异性和菌株特异性。例如,Federici等[20]研究了双歧杆菌属内某些菌株的草酸盐降解活性,发现动物双歧杆菌乳亚种DSM 10140的草酸盐降解活性最高,能降解培养基中61%的草酸盐;而长双歧杆菌MB 282和青春双歧杆菌MB 238的草酸盐降解率分别为35%和57%,同为短双歧杆菌的MB 283菌株可降解38%的草酸盐,而MB 151菌株的草酸盐降解率仅为1%;此外,供试的5 株长双歧杆菌中仅有4 株能降解草酸盐。
表1 具有降解草酸盐活性乳酸菌的主要种属
Table 1 Major genera and species of lactic acid bacteria with oxalate-degrading activity
2 乳酸菌降解草酸盐的特性
2.1 体外实验
体外实验通常作为筛选具有益生功能乳酸菌的首要步骤。研究乳酸菌草酸盐降解活性的实验是将供试乳酸菌菌株培养在含草酸盐的培养基中,通过培养前后培养基中草酸盐含量变化计算降解率进而评价其草酸盐降解活性。2000年,Hokama等[13]首次报道了乳酸菌具有降解草酸盐的能力,分离到11 株可降解草酸盐的粪肠球菌,其中有4 株菌培养2 d后,其培养基中草酸盐(2.0 g/L)全部被降解,同时发现粪肠球菌需要一种贫瘠的营养生境且重复传代培养才能维持其降解草酸盐的活性。随后,研究人员通过体外实验筛选到许多其他种属的乳酸菌也具有草酸盐降解活性,如广泛用于乳制品和药品的12 株双歧杆菌在含有5 mmol/L草酸钠的培养基中培养5 d,供试菌株中动物双歧杆菌乳亚种DSM 10140菌株草酸盐降解活性最强,可降解培养基中61%的草酸钠[20]。Weese等[21]对来自犬粪便样品的37 株乳酸菌进行体外降解草酸盐活性研究,发现降解率为0%~65%,其中4 株菌无草酸盐降解活性。同样来源于犬粪便的47 株乳酸菌中,49%菌株无草酸盐降解活性,34%菌株对草酸盐的降解率低于5%[31]。13 株乳杆菌和5 株双歧杆菌在含10 mmol/L草酸铵的培养基中培养24 h后,发现供试乳杆菌的草酸盐降解活性高于双歧杆菌[29]。由此说明,乳酸菌降解草酸盐的能力具有菌株特异性。同时有研究发现,乳酸菌降解草酸盐的能力明显低于食草酸杆菌,食草酸杆菌几乎可100%降解培养基中的草酸盐[20,29]。为了更接近人体内真实情况,研究人员通过模拟人体结肠微生态条件评价了格氏乳杆菌Gasser AM63T的草酸盐降解活性,发现其在构建的三阶段连续培养系统的第一个阶段(即近端结肠)就出现了草酸盐降解活性[22]。此外,甲酰辅酶A转移酶(formyl-CoA transferase,Frc)和草酰辅酶A脱羧酶(oxalyl-CoA decarboxylase,Oxc)作为人体内草酸盐降解的2 个关键酶,其编码基因和功能在乳酸菌中已得以鉴定[13,20,25,35]。因此,具有Frc基因frc和Oxc基因oxc一度成为体外分离筛选可降解草酸盐乳酸菌菌株的依据之一[26]。
2.2 体内实验
动物模型数据尤其是人体临床实验结果能最直接证明乳酸菌益生功能在体内能否真正发挥作用。目前已有大量研究证实了乳酸菌在体内具有抑制肠道草酸盐吸收的作用。
高草酸尿症及相关肾脏疾病实验的模型动物通常以大鼠、小鼠以及犬最为常见。Kwak等[28]研究了乳杆菌对大鼠尿路结石形成的预防作用,与对照组相比,体外具有降解草酸盐活性的干酪乳杆菌HY2743和HY7201菌株可明显降低大鼠尿草酸的分泌,并且可使模型大鼠肾脏内晶体形成量明显减少。Klimesova等[36]研究了一株双歧杆菌对原发性高草酸尿症小鼠模型尿草酸分泌的影响,发现动物双歧杆菌乳亚种DSM 10140可降低模型小鼠尿草酸分泌,不同于食草酸杆菌通过促进肠道草酸分泌来降低尿草酸的分泌量[37],这种作用是该菌株通过降解饮食草酸盐从而限制肠道对草酸盐的吸收来完成的。Gomathi等[16]以富含草酸钾的饮食诱导大鼠构建高草酸尿和草酸钙结石模型,以体外具有降解草酸盐活性的乳酸菌为受试对象,发现5×108CFU/mL剂量的供试发酵乳杆菌AB1、TY5菌株和唾液乳杆菌AB11菌株组成的三联菌(活菌数之比为1∶1∶1)灌胃给予模型大鼠21 d后,模型大鼠的尿草酸水平明显降低。钟文婷[32]研究了一株乳酸乳球菌对食源性草酸过多导致的犬尿石症的预防作用,发现在乳酸乳球菌处理第15天时模型犬尿草酸分泌量降低了21%,但差异不显著,而在处理第30天时模型犬尿草酸分泌量下降了33%,且差异显著(P<0.05),同时发现处理30 d后,犬尿钙含量降低了10%,且差异显著(P<0.05);说明该菌株具有潜在的降低尿草酸和尿钙分泌的能力。
乳酸菌的草酸盐降解活性在人体临床实验上也取得了较好的效果,尤以商业化的可降解草酸盐的乳酸菌产品Oxadrop®和VSL#3®研究报道最多[33-34,38-39]。Campieri等[38]以原发性草酸尿路结石症或弱高草酸尿症患者为研究对象,每次给予活菌数为8×1011个的冻干乳酸菌混合物Oxadrop®(嗜酸乳杆菌、植物乳杆菌、短乳杆菌、嗜热链球菌和婴儿双歧杆菌的活菌数之比为1∶1∶1∶1∶1),每天2 次,重悬于冷水中,饭前服用,连续服用4 周后,患者24 h的尿草酸分泌量从55.5 mg降到33.5 mg,下降了40%,实验结束1 个月后,尿草酸分泌量继续下降到28.3 mg,下降了49%。随后,Lieske等[33]同样以Oxadrop®为研究对象,考察了其对肠源性高草酸尿症人群的影响,但使用的Oxadrop®产品菌种组成及比例(嗜酸乳杆菌、短乳杆菌、嗜热链球菌和婴儿双歧杆菌活菌数之比为1∶1∶4∶4)与Campieri等[38]所使用的略有不同,受试人群每天服用1 次该产品(活菌数8×1011个),服用1 个月后尿草酸分泌量下降了19%,第2个月改为每天服用2次,尿草酸分泌量下降了24%,第3个月改为每天服用3 次,尿草酸分泌量相比于初始值仅下降了2%,而且停药1 个月后,尿草酸分泌量仍下降了20%,这说明乳酸菌在人体肠道内发生定植并继续发挥作用。Okombo等[39]研究了VSL#3®产品对健康人体消化道草酸盐吸收的影响,VSL#3®产品由嗜热链球菌、短双歧杆菌、长双歧杆菌、婴儿双歧杆菌、嗜酸乳杆菌、植物乳杆菌、副干酪乳杆菌、德氏乳杆菌保加利亚亚种8 种活性乳酸菌组成,受试个体每天摄取含相同量的低草酸盐的食物,并口服80 mg草酸盐,同时给予VSL#3®产品(8×1011个活菌),发现服用产品4 周后,总草酸盐在22 h内的吸收率从30.8%降到11.6%,并且停用产品4 周后,22 h内的吸收率仍能维持在11.5%水平,但对于6 h内的吸收率,与服用4 周后、服用前均无显著差异;由此推测,VSL#3®产品对草酸盐的降解作用可能发生在结肠部位。Al-Wahsh等[34]考察了VSL#3®产品对人体消化道对草酸盐吸收的影响,以11 名健康个体为研究对象,对照组口服草酸钠,低剂量组和高剂量组除口服草酸钠外分别口服1 粒和2 粒VSL#3®胶囊,每粒VSL#3®胶囊中含有4.5×1011个活菌,结果发现,该产品可明显降低受试个体尿草酸水平,但高低两个剂量组在草酸盐吸收上没有明显差异。然而,Goldfarb等[40]在一项随机控制临床实验中,考察了Oxadrop®产品对原发性高草酸尿症人群的影响,与Lieske等[33]研究中使用Oxadrop®产品相比,该临床实验所用的Oxadrop®产品的菌种组成和比例不变,患者每天服用剂量(活菌数1.08×1012个)略高但无实质性差异,服用该产品4 周后,患者在24 h内的尿草酸分泌量虽然降低了6%,但与服用前的差异并不显著,与此类似的结果也出现在Siener等[41]的研究报道中。由此说明,Oxadrop®产品在人体临床实验报道中并不都是积极的结果,这可能是源于人体临床实验相比动物实验,在饮食上往往难于控制,同时受试个体数量有限且个体之间差异较大。
3 乳酸菌降解草酸盐的机理
目前,草酸盐降解菌研究最为清楚的是食草酸杆菌,草酸盐是其生长代谢的唯一或主要碳源,因此被称为“专性草酸营养型”[42],其对草酸盐的降解首先是利用草酸盐-甲酸盐反向转运体OxlT(oxlT基因编码)将胞外草酸盐吸收到细胞内,然后再利用Frc(frc基因编码)将胞内草酸盐转化成草酰辅酶A,草酰辅酶A再通过Oxc(oxc基因编码)的作用脱羧形成甲酸盐和CO2,最后再利用OxlT将胞内甲酸盐运输到胞外,从而完成对草酸盐的降解作用[43]。
与食草酸杆菌不同,乳酸菌属于兼性草酸营养型微生物,即不以草酸盐作为其生长的主要碳源,仅能利用草酸盐作为碳源[38]。乳酸菌对草酸盐的降解机理与食草酸杆菌类似,最早是在一株粪肠球菌中被阐述[13]。研究人员通过蛋白质免疫印迹技术检测到可降解草酸盐的粪肠球菌中存在2 种蛋白,可以同抗食草酸杆菌源的Frc和Oxc蛋白的抗体发生免疫反应;由此说明,粪肠球菌中降解草酸盐的酶类与食草酸杆菌中的酶相似[13],但并没有发现草酸盐转运蛋白的存在,粪肠球菌降解草酸盐的确切机制尚未完全明确。Federici等[20]以已公布的食草酸杆菌oxc基因为模板设计引物,在一株具有草酸盐降解活性的动物双歧杆菌乳亚种中通过PCR反应成功扩增出同源体,其氨基酸序列与食草酸杆菌Oxc的氨基酸序列同源性为47%,在大肠杆菌中过表达的蛋白能够与抗食草酸杆菌的Oxc抗体发生杂交,并通过酶活性分析证实了动物双歧杆菌乳亚种源oxc基因的Oxc功能活性。随后,Turroni等[30]在动物双歧杆菌乳亚种中鉴定出了oxc和frc基因,细胞预适应于亚抑制浓度草酸盐条件后,将其暴露于pH 4.5条件下,发现其草酸盐降解率显著提高,证实了酸性条件是动物双歧杆菌乳亚种产生较高草酸盐降解率的前提。乳杆菌属的草酸盐降解机制同样是围绕Frc和Oxc提出,Azcarate-Peril等[25]在嗜酸乳杆菌NCFM中鉴定了编码降解草酸盐的Frc和Oxc的基因,发现这两个基因只有在细菌细胞适应亚抑制浓度草酸盐并且在弱酸性条件下(pH 5.5)才能发挥功能。在格氏乳杆菌Gasser AM63T中,同样检测到了frc和oxc基因,也需要在弱酸性、同时有草酸盐存在的条件下才能够被诱导表达[22]。Turroni等[26]发现草酸盐降解率超过50%的所有供试嗜酸乳杆菌和格氏乳杆菌中均存在oxc和frc基因。Kullin等[44]对一株罗伊氏乳杆菌100-23C进行全基因组分析,发现其基因组中存在紧邻的oxc和frc基因,预测的Oxc和Frc产物分别与格氏乳杆菌、嗜酸乳杆菌和动物双歧杆菌乳亚种中的Oxc和Frc具有高度的同源性,但该菌株野生型并不能降解培养基中的草酸盐,这可能是由于其缺少草酸盐转运体。由此说明,细菌的基因组中含有oxc和frc基因并不足以证明其一定具有降解草酸盐的能力。除了Oxc和Frc之外,还需草酸盐转运体将草酸盐由胞外转移到胞内,而Turroni等[30]在动物双歧杆菌乳亚种中发现了一个可能与草酸盐转运有关的编码透性酶的开放阅读框,并且存在于已公布的全基因组序列中,这种透性酶基因也存在于许多可降解草酸盐的乳杆菌属菌株中;然而,在罗伊氏乳杆菌100-23C全基因组中并不存在该透性酶基因,这可能是该菌株不能降解草酸盐的主要原因。
综上所述,乳酸菌通过透性酶将胞外草酸盐转运到胞内,通过Frc将草酸盐转化为草酰辅酶A,再通过Oxc脱羧产生甲酸盐和CO2完成对草酸盐的降解。
4 结 语
不论是体外实验还是体内实验结果均表明,乳酸菌的草酸盐降解活性无法与食草酸杆菌相媲美。食草酸杆菌的生长需要以草酸盐作为其生长代谢的唯一或主要碳源,因此,食草酸杆菌对治疗高草酸尿症非常有效。此外,肠道内草酸盐浓度的高低受饮食组成影响较大,这也是食草酸杆菌在肠道内定植不稳定的根本原因。相比之下,乳酸菌能够利用草酸盐但不以其作为主要碳源,在肠道内的定植不受肠道草酸盐水平的限制,可稳定定植于肠道内。因此,乳酸菌在预防高草酸尿症方面具有独特的优势。可降解草酸盐的乳酸菌主要分布于乳杆菌属、双歧杆菌属以及肠球菌属,且具有菌种和菌株特异性,其降解草酸盐的作用受3个基因调控,即oxc、frc及编码透性酶的基因,但关于透性酶的遗传信息及发挥功能的确切机制仍需更深入的研究加以证实。总之,乳酸菌的草酸盐降解活性是明确的,在预防与治疗草酸盐相关的肾脏疾病上具有较好的应用前景。
参考文献:
[1] COCHAT P, RUMSBY G. Primary hyperoxaluria[J]. The New England Journal of Medicine, 2013, 369: 649-658. DOI:10.1056/NEJMra1301564.
[2] BHASIN B, ÜREKLI H M, ATTA M G. Primary and secondary hyperoxaluria: understanding the enigma[J]. World Journal of Nephrology, 2015, 4(2): 235-244. DOI:10.5527/wjn.v4.i2.235.
[3] WEINMAN E J, FRANKFURT S J, INCE A, et al. Renal tubular transport of organic acids. Studies with oxalate and para-aminohippurate in the rat[J]. The Journal of Clinical Investigation, 1978, 61(3): 801-806.DOI:10.1172/JCI108994.
[4] STEF R, BERND H, BENJAMIN A V, et al. Hyperoxaluria: a gut-kidney axis?[J]. Kidney International, 2011, 80(11): 1146-1158.DOI:10.1038/ki.2011.287.
[5] KHODOR S A, SHATAT I F. Gut microbiome and kidney disease:a bidirectional relationship[J]. Pediatric Nephrology, 2016, 32(6): 921-931.DOI:10.1007/s00467-016-3392-7.
[6] JAIRATH A, PAREKH N, OTANO N, et al. Oxalobacter formigenes: opening the door to probiotic therapy for the treatment of hyperoxaluria[J]. Scandinavian Journal of Urology and Nephrology, 2015,49(4): 334-337. DOI:10.3109/21681805.2014.996251.
[7] ELLIS M L, SHAW K J, JACKSON S B, et al. Analysis of commercial kidney stone probiotic supplements[J]. Urology, 2015, 85(3): 517-521.DOI:10.1016/j.urology.2014.11.013.
[8] ALLISON M J, DAWSON K A, MAYBERRY W R, et al. Oxalobacter formigenes gen. nov., sp. nov.: oxalate-degrading anaerobes that inhabit the gastrointestinal tract[J]. Archives of Microbiology, 1985, 141(1): 1-7.DOI:10.1007/BF00446731.
[9] JIANG J, KNIGHT J, EASTER L H, et al. Impact of dietary calcium and oxalate, and Oxalobacter formigenes colonization on urinary oxalate excretion[J]. Journal of Urology, 2011, 186(1): 135-139. DOI:10.1016/j.juro.2011.03.006.
[10] LI X S, ELLIS M L, KNIGHT J. Oxalobacter formigenes colonization and oxalate dynamics in a mouse model[J]. Applied and Environmental Microbiology, 2015, 81(15): 5048-5054. DOI:10.1128/AEM.01313-15.
[11] BARNETT C, NAZZAL L, GOLDFARB D S, et al. The presence of Oxalobacter formigenes in the microbiome of healthy young adults[J]. Journal of Urology, 2016, 195(2): 499-506. DOI:10.1016/j.juro.2015.08.070.
[12] LANGE J N, WOOD K D, WONG H, et al. Sensitivity of human strains of Oxalobacter formigenes to commonly prescribed antibiotics[J]. Urology,2012, 79(6): 1286-1289. DOI:10.1016/j.urology.2011.11.017.
[13] HOKAMA S, HONMA Y, TOMA C, et al. Oxalate-degrading Enterococcus faecalis[J]. Microbiology and Immunology, 2000, 44(4):235-240. DOI:10.1111/j.1348-0421.2000.tb02489.x.
[14] FERRAZ R R, MARQUES N C, FROEDER L, et al. Effects of Lactobacillus casei and Bifidobacterium breve on urinary oxalate excretion in nephrolithiasis patients[J]. Urological Research, 2009, 37(2):95-100. DOI:10.1007/s00240-009-0177-5.
[15] GIARDINA S, SCILIRONI C, MICHELOTTI A, et al. In vitro antiinflammatory activity of selected oxalate-degrading probiotic bacteria:potential applications in the prevention and treatment of hyperoxaluria[J].Journal of Food Science, 2014, 79(3): M384-M390. DOI:10.1111/1750-3841.12344.
[16] GOMATHI S, SASIKUMAR P, ANBAZHAGAN K, et al. Oral administration of indigenous oxalate degrading lactic acid bacteria and quercetin prevents calcium oxalate stone formation in rats fed with oxalate rich diet[J]. Journal of Functional Foods, 2015, 17: 43-54.DOI:10.1016/j.jff.2015.05.011.
[17] PECK A B, CANALES B K, NGUYEN C Q. Oxalate-degrading microorganisms or oxalate-degrading enzymes: which is the future therapy for enzymatic dissolution of calcium-oxalate uroliths in recurrent stone disease?[J] Urolithiasis, 2016, 44(1): 45-50. DOI:10.1007/s00240-015-0845-6.
[18] DI CERBO A, PEZZUTO F, PALMIERI L, et al. Clinical and experimental use of probiotic formulations for management of end-stage renal disease: an update[J]. International Urology and Nephrology, 2013,45(6): 1569-1576. DOI:10.1007/s11255-012-0335-5.
[19] 任志华. 草酸降解菌的分离鉴定及其对犬草酸钙尿结石的预防作用与机理研究[D]. 南京: 南京农业大学, 2011: 115-123.
[20] FEDERICI F, VITALI B, GOTTI R, et al. Characterization and heterologous expression of the oxalyl coenzyme A decarboxylase gene from Bifidobacterium lactis[J]. Applied and Environmental Microbiology, 2004, 70(9): 5066-5073. DOI:10.1128/AEM.70.9.5066-5073.2004.
[21] WEESE J S, WEESE H E, YURICEK L, et al. Oxalate degradation by intestinal lactic acid bacteria in dogs and cats[J]. Veterinary Microbiology,2004, 101(3): 161-166. DOI:10.1016/j.vetmic.2004.03.017.
[22] LEWANIKA T R, REID S J, ABRATT V R, et al. Lactobacillus gasseri Gasser AM63Tdegrades oxalate in a multistage continuous culture simulator of thehuman colonic microbiota[J]. FEMS Microbiology Ecology, 2007, 61(1): 110-120. DOI:10.1111/j.1574-6941.2007.00327.x.
[23] GOMATHI S, SASIKUMAR P, ANBAZHAGAN K, et al. Screening of indigenous oxalate degrading lactic acid bacteria from human faeces and South Indian fermented foods: assessment of probiotic potential[J]. The Scientiベc World Journal, 2014, 2014: 648059. DOI:10.1155/2014/648059.
[24] 董婷婷. 降解草酸益生菌的筛选及其对实验动物的功效研究[D]. 大连: 大连医科大学, 2012: 6-28.
[25] AZCARATE-PERIL M A, BRUNO-BÁRCENA J M, HASSAN H M,et al. Transcriptional and functional analysis of oxalyl-coenzyme A (CoA)decarboxylase and formyl-CoA transferase genes from Lactobacillus acidophilus[J]. Applied and Environmental Microbiology, 2006, 72(3):1891-1899. DOI:10.1128/AEM.72.3.1891-1899.2006.
[26] TURRONI S, VITALI B, BENDAZZOLI C, et al. Oxalate consumption by lactobacilli evaluation of oxalyl-CoA decarboxylase and formyl-CoA transferase activity in Lactobacillus acidophilus[J]. Journal of Applied Microbiology, 2007, 103(5): 1600-1609. DOI:10.1111/j.1365-2672.2007.03388.x.
[27] CHO J G, GEBHART C J, FURROW E, et al. Assessment of in vitro oxalate degradation by Lactobacillus species cultured from veterinary probiotics[J]. American Journal of Veterinary Research, 2015, 76(9): 801-806. DOI:10.2460/ajvr.76.9.801.
[28] KWAK C, JEONG B C, KU J H, et al. Prevention of nephrolithiasis by Lactobacillus in stone-forming rats: a preliminary study[J]. Urological Research, 2006, 34(4): 265-270. DOI:10.1007/s00240-006-0054-4.
[29] MOGNA L, PANE M, NICOLA S, et al. Screening of different probiotic strains for their in vitro ability to metabolise oxalates: any prospective use in humans?[J] Journal of Clinical Gastroenterology, 2014, 48: S91-S95.DOI:10.1097/MCG.0000000000000228.
[30] TURRONI S, BENDAZZOLI C, DIPALO S C F, et al. Oxalatedegrading activity in Biベdobacterium animalis subsp. lactis: impact of acidic conditions on the transcriptional levels of the oxalyl coenzyme A(CoA) decarboxylase and formyl-CoA transferase genes[J]. Applied and Environmental Microbiology, 2010, 76(16): 5609-5620. DOI:10.1128/AEM.00844-10.
[31] REN Z, PAN C, JIANG L, et al. Oxalate-degrading capacities of lactic acid bacteria in canine feces[J]. Veterinary Microbiology, 2011, 152(3/4):368-373. DOI:10.1016/j.vetmic.2011.05.003.
[32] 钟文婷. 富硒乳酸菌对食源性草酸过多犬尿石症的预防作用[D]. 南京: 南京农业大学, 2013: 31-43.
[33] LIESKE J C, GOLDFARB D S, DE SIMONE C, et al. Use of a probiotic to decrease enteric hyperoxaluria[J]. Kidney International, 2005,68(3): 1244-1249. DOI:10.1111/j.1523-1755.2005.00520.x.
[34] AL-WAHSH I, WU Y, LIEBMAN M. Acute probiotic ingestion reduces gastrointestinal oxalate absorption in healthy subjects[J]. Urological Research, 2012, 40(3): 191-196. DOI:10.1007/s00240-011-0421-7.
[35] ALTERMANN E, RUSSELL W M, AZCARATE-PERIL M A, et al. Complete genome sequence of the probiotic lactic acid bacterium Lactobacillus acidophilus NCFM[J]. Proceedings of the National Academy of Sciences of the United States of Aemrica, 2005, 102(11):3906-3912. DOI:10.1073/pnas.0409188102.
[36] KLIMESOVA K, WHITTAMORE J M, HATCH M. Biベdobacterium animalis subsp. lactis decreases urinary oxalate excretion in a mouse model of primary hyperoxaluria[J]. Urolithiasis, 2015, 43(2): 107-117.DOI:10.1007/s00240-014-0728-2.
[37] HATCH M, CORNELIUS J, ALLISON M, et al. Oxalobacter sp. reduces urinary oxalate excretion by promoting enteric oxalate secretion[J]. Kidney International, 2006, 69(4): 691-698. DOI:10.1038/sj.ki.5000162.
[38] CAMPIERI C, CAMPIERI M, BERTUZZI V, et al. Reduction of oxaluria after an oral course of lactic acid bacteria at high concentration[J].Kidney International, 2001, 60(3): 1097-1105. DOI:10.1046/j.1523-1755.2001.0600031097.x.
[39] OKOMBO J, LIEBMAN M. Probiotic-induced reduction of gastrointestinal oxalate absorption in healthy subjects[J]. Urological Research, 2010, 38(3): 169-178. DOI:10.1007/s00240-010-0262-9.
[40] GOLDFARB D S, MODERSITZKI F, ASPLIN J R. A randomized,controlled trial of lactic acid bacteria for idiopathic hyperoxaluria[J].Clinical Journal of the American Society of Nephrology, 2007, 2(4): 745-749. DOI:10.2215/CJN.00600207.
[41] SIENER R, BADE D J, HESSE A, et al. Dietary hyperoxaluria is not reduced by treatment with lactic acid bacteria[J]. Journal of Translational Medicine, 2013, 11: 306. DOI:10.1186/1479-5876-11-306.
[42] SAHIN N. Oxalotrophic bacteria[J]. Research in Microbiology, 2003,154(6): 399-407. DOI:10.1016/S0923-2508(03)00112-8.
[43] ABRATT V R, REID S J. Oxalate-degrading bacteria of the human gut as probiotics in the management of kidney stone disease[J]. Advances in Applied Microbiology, 2010, 72: 63-87. DOI:10.1016/S0065-2164(10)72003-7.
[44] KULLIN B, TANNOCK G W, LOACH D M, et al. A functional analysis of the formyl-coenzyme A (frc) gene from Lactobacillus reuteri 100-23C[J]. Journal of Applied Microbiology, 2014, 116(6): 1657-1667.DOI:10.1111/jam.12500.