
图1 不同CaCl2质量浓度对C. oleophila生长情况的影响
Fig. 1 Effects of different concentrations of CaCl2 on the growth of C. oleophila
果蔬采后生物防治关键在于拮抗微生物能够在果皮或体内定植,通过自身的不断繁殖从而对病原菌产生对抗作用,而酵母和病原真菌都属于异养生物,必须从外界吸收营养物质来维持生存。经研究发现,橄榄假丝酵母(Candida oleophila)可能的抑菌机制为营养和空间的竞争,当拮抗菌数量充足时,拮抗酵母能够抢占更多的营养物质和空间,从而抑制病原菌的生长[1-2]。现阶段的研究发现,Pichia caribbica、Pichia guilliermondii、Aureobasidium pullulans等拮抗菌均可以通过营养与空间竞争的作用机制减少果蔬采后真菌病害[3-5]。当向酵母添加外源营养物时,营养物的种类、添加量都会对酵母的控制病害效果产生影响。目前有研究表明,拮抗微生物通过结合某些佐剂即营养物质(如某一种糖类、氨基酸等)可以增加其群体数量并增强其存活率,同时,病原菌对这些佐剂的利用率则较低,在两种因素共同作用下,控病效果大大提升。除了营养和空间的竞争机制外,诱导果实自身产生抗病性也是拮抗酵母作用的机制之一[6-7]。当植物体受到生物、化学或者物理因素影响时,会刺激其自身抵抗病原菌的相关防御系统,酵母作为一种生物激发子,可以诱导植物体自身产生一系列抗性相关的物质[8-9]。拮抗酵母产生的诱导抗病性可以促进植物组织产生病程相关蛋白家族和酚类代谢系统中的一些酶如过氧化物酶、苯丙氨酸解氨酶、几丁质酶(chitinase,CHI)、β-1,3-葡聚糖酶(β-1,3-glucanase,GLU)的活力,以及促进木质素和酚类物质的生成等[7,10]。
近几年的研究结果表明,CaCl2可以和多种酵母结合使用,增强拮抗酵母菌对多种果实病害的生防效力。然而,有关CaCl2和C. oleophila结合使用控制苹果果实采后青霉病病害的相关研究却鲜见报道。本研究先通过离体实验考察CaCl2对C. oleophila和扩展青霉菌(Penicillium expansum)生长状况的影响,在此基础上进行苹果果实活体实验,筛选最佳的复合比例并探究最佳复合质量浓度CaCl2结合C. oleophila处理果实,增强C. oleophila对苹果果实采后青霉病的控制效果。
‘红富士’苹果产自甘肃静宁,取大小均一、无机械伤、成熟度基本一致的果实作为实验材料。经质量分数为2%的次氯酸钠浸泡2 min后,自来水冲洗,晾干备用[11]。
C. oleophila分离自重庆北碚歇马镇冯家槽柑桔树(‘锦橙447’)叶片表面。该菌株在NYDA培养基(酵母膏5 g、牛肉膏8 g、葡萄糖10 g、琼脂20 g、水1 000 mL)中、4 ℃条件下保藏。每两个月活化一次。
扩展青霉菌分离自发病苹果果实。采用组织分离法从发病苹果果实上分离病原菌,且致病性检测结果、病原菌菌落形态、菌丝形态以及分子生物学鉴定结果表明其为扩展青霉菌。
SW-CJ-1F超净工作台 苏净集团安泰有限公司;B203生物显微镜 重庆奥特光学仪器有限公司;BS-4G振荡培养箱 金坛市富华仪器有限公司;DHP-9082电热恒温培养箱 上海齐欣科学仪器有限公司;PSX-280手提式高压杀菌锅 上海申安医疗器械厂;XB.K.25血球计数板型 上海求精生化试剂有限公司;GL-20G-Ⅱ高速冷冻离心机 上海安亭科学仪器厂;SL602N高精显电子天平 上海民桥精密科学仪器有限公司;LGJ-10真空冷冻干燥机 北京松源华兴有限公司。
1.3.1 离体条件下CaCl2对C. oleophila和P. expansum生长状况的影响
1.3.1.1 CaCl2在NYDB培养基中对C. oleophila生长状况的影响
在含90 mL NYDB液体培养基的试管中分别加入10 mL不同质量浓度的CaCl2溶液,使试管中CaCl2的最终质量浓度为0、10、20、50、100 g/L。将2 mL 1×108 CFU/mL的C. oleophila细胞悬浮液接种于液体培养基内,在28 ℃、200 r/min的摇床中培养24 h后计数。每个处理组重复3 次。
1.3.1.2 CaCl2在PDA培养基中对P. expansum生长状况的影响
用移液器吸取1 μL 1×106 CFU/mL的P. expansum孢子悬浮液点在含有不同质量浓度CaCl2(0、5、10、20 g/L)PDA培养基中央。25 ℃条件下培养7 d后拍照。每个处理组重复3 次。
1.3.2 CaCl2与C. oleophila复合处理对苹果果实采后青霉病的抑制作用
1.3.2.1 CaCl2与C. oleophila复合处理对苹果果实采后青霉病的抑制效果
将果实随机分成6 组。用无菌打孔器在果实赤道部位等距离打孔(直径3 mm,深度3 mm),每个苹果打3 个伤口,每个处理组果实伤口处分别加入20 μL下述液体:1)无菌水(阳性对照);2)1×107 CFU/mL C. oleophila悬浮液;3)5 g/L CaCl2与1×107 CFU/mL C. oleophila混合悬浮液;4)10 g/L CaCl2与1×107 CFU/mL C. oleophila混合悬浮液;5)20 g/L CaCl2与1×107 CFU/mL C. Oleophila混合悬浮液;6)50 g/L CaCl2与1×107 CFU/mL C. oleophila混合悬浮液。4 h后,在果实伤口处接入20 μL的1×105 CFU/mL P. expansum孢子悬浮液,待菌液吸收后,单果包装,置于25 ℃、相对湿度90%~95%条件下贮藏,定期统计果实发病率和病斑直径。每个处理组10 个果实,重复3 次。
1.3.2.2 CaCl2复合处理对C. oleophila在苹果果实伤口生长动态的影响
伤口接种处理:将果实随机分成2 组。用无菌打孔器在果实赤道部位等距离地打伤口(直径3 mm,深度3 mm),每个苹果打3 个伤口,每个伤口处分别加入20 μL的1×107 CFU/mL C. oleophila悬浮液、10 g/L CaCl2与1×107 CFU/mL C. oleophila混合悬浮液。待菌液吸收后,单果包装,在25 ℃、相对湿度90%~95%环境下贮藏,以接种后1 h测定的酵母菌数为起始值,每天取一次样,用稀释平板法测定酵母数目。取样时,用消毒的打孔器取伤口处直径为1 cm的果肉组织10 份,放入含10 mL无菌水的消毒研钵内研磨至匀浆,25 ℃条件下培养48 h后计数,结果以每伤口处酵母数量为单位(lg(cells/wound)),每个处理组重复3 次,每个重复10 个果实,实验重复3 次。
1.3.3 CaCl2与C. oleophila复合处理诱导苹果果实抗病性机制
1.3.3.1 样品处理及取样方法
将果实随机分成3 组。用无菌打孔器在果实赤道部位等距离地打伤口(直径3 mm,深度3 mm),每个苹果打3 个伤口,每个伤口处分别加入20 μL的无菌水(阳性对照)、1×107 CFU/mL C. oleophila悬浮液、10 g/L CaCl2与1×107 CFU/mL C. oleophila混合悬浮液。贮藏及取样方法同1.3.2.2节,每个处理组重复3 次,每个重复30 个果实,实验重复3 次。
1.3.3.2 CHI、GLU活力测定
酶提取液制备:称取1.0 g苹果果肉,加入4 mL 200 mmol/L pH 5.2的醋酸缓冲液,在冰浴条件下研磨匀浆,于4 ℃、8 000 r/min离心30 min,收集上清液用于CHI和GLU活力测定。
CHI活力的测定参考Boller等[12]的方法并略作修改。取0.05 mL粗酶液,加入0.5 mL 10 mg/mL胶状几丁质悬浮液和0.95 mL 100 mmol/L pH 5.2的醋酸缓冲液,混合后在37 ℃保温培养1 h。然后往该混合液中加入0.1 mL、质量分数为3%的脱盐蜗牛酶,继续37 ℃保温培养70 min,立即加入0.2 mL 0.6 mol/L的四硼酸钾溶液,沸水浴3 min,然后迅速冷却。冷却完毕,加入2 mL用冰醋酸稀释5 倍体积的对二甲氨基苯甲醛储备液。再37 ℃保温培养30 min显色,然后测定585 nm波长处的吸光度。以煮沸5 min酶液作对照,根据标准曲线,计算N-乙酰葡萄糖胺(Glc-Nac)生成量,以每秒钟每克蛋白分解胶状几丁质产生1×10-9 mol Glc-NAc为一个酶活力单位(U)。实验重复3 次。
GLU活力测定参考Abeles等[13]的方法并略作修改。取100 µL粗酶液,加入100 µL质量分数0.4%昆布多糖溶液,于37 ℃保温反应45 min后,加入1.8 mL蒸馏水和1.5 mL DNS试剂,沸水浴加热5 min终止酶促反应。煮沸后迅速冷却,然后用蒸馏水将显色的反应液稀释至25 mL,混匀,测定540 nm波长处的吸光度。以加热煮沸5 min的酶液作对照,根据标准曲线,计算生成还原糖的量,以每秒钟每毫克蛋白分解昆布多糖产生1×10-9 mol葡萄糖为一个酶活力单位(U)。实验重复3 次。
1.3.3.3 木质素含量的测定
木质素含量的测定参照Li Hua等[14]的方法并略作修改。称取1 g样品,加5 mL、体积分数95%乙醇溶液研磨,经8 000 r/min离心10 min,沉淀物以3 mL 95%乙醇溶液冲洗3 次,再用3 mL乙醇-正己烷(1∶2,V/V)离心冲洗3 次,收集沉淀物,于65 ℃环境下干燥12 h左右。干燥物溶于3 mL、体积分数25%溴乙酰冰醋酸溶液中,在70 ℃恒温水浴中加塞保温30 min,然后加0.9 mL 2 mol/L NaOH溶液终止反应,再以冰醋酸定容至10 mL,8 000 r/min离心10 min,吸取上清液0.50 mL加入到9.50 mL蒸馏水中,以蒸馏水为参比调零,在280 nm波长处测定光密度。以每克鲜质量样品在280 nm处的光密度(OD值)表示木质素含量。重复3 次。
1.3.3.4 总酚和类黄酮含量的测定
总酚和类黄酮含量测定参照Deng Lili等[15]的方法进行测定。1.0 g样品与预冷的20 mL、体积分数1%HCl-甲醇溶液充分研磨提取,然后4 ℃、12 000×g离心10 min,上清液直接用于比色。总酚含量以每克鲜质量样品在280 nm波长处的光密度(OD值)表示,类黄酮含量以每克鲜质量样品在325 nm波长处的光密度(OD值)表示。实验重复3 次。
采用Excel 2016软件统计分析所有数据,计算标准差并制图;应用SPSS 22软件进行方差分析,利用邓肯氏多重比较对差异显著性进行分析,P<0.05表示差异显著。
2.1.1 CaCl2在NYDB培养基中对C. oleophila生长状况的影响
CaCl2在NYDB培养基中对C. oleophila生长状况的影响如图1所示,在28 ℃下培养24 h后,当CaCl2质量浓度小于50 g/L时,CaCl2对C. oleophila生长有显著促进作用(P<0.05);100 g/L CaCl2对C. oleophila生长有显著抑制作用(P<0.05)。说明不同质量浓度的CaCl2对C. oleophila的生长状况有不同影响。
图1 不同CaCl2质量浓度对C. oleophila生长情况的影响
Fig. 1 Effects of different concentrations of CaCl2 on the growth of C. oleophila
2.1.2 CaCl2在PDA培养基中对P. expansum生长状况的影响
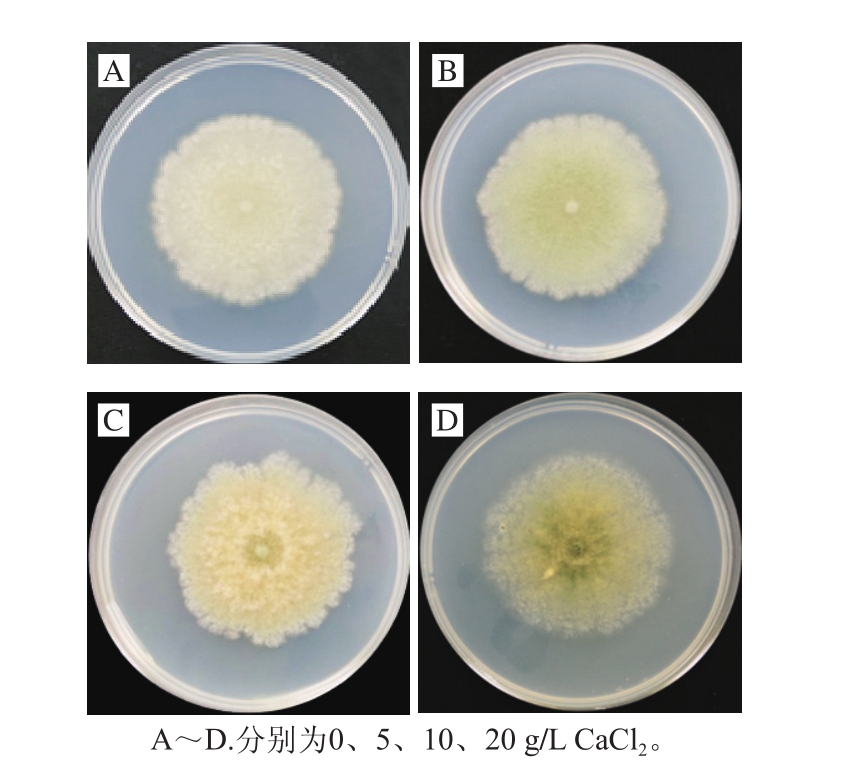
图2 不同质量分数CaCl2对P. expansum生长状况的影响
Fig. 2 Inhibitory effects of different concentrations of calcium chloride on mycelial growth of P. expansum
CaCl2在PDA培养基中对P. expansum生长状况的影响如图2所示,在25 ℃下培养7 d后,与对照组(0 g/L)相比,5 g/L CaCl2处理对P. expansum生长有促进作用,其P. expansum产孢能力较强。与对照组相比,10、20 g/L CaCl2处理对P. expansum生长有抑制作用,其P. expansum产孢能力弱。说明不同质量浓度的CaCl2对P. expansum的生长状况有不同影响。
2.2.1 CaCl2与C. oleophila复合处理对苹果果实采后青霉病的抑制效果
CaCl2与C. oleophila复合使用对苹果果实采后青霉病的抑制效果如图3所示,在贮藏3 d和4 d时,5 个处理组果实的发病率和病斑直径显著低于对照果实(P<0.05),说明这5 种处理均能有效延缓苹果果实采后青霉病的发生。在整个贮藏期间,C. oleophila+10 g/L CaCl2处理组果实的发病率和病斑直径显著低于其他5 组(P<0.05),说明其控制病害的效果最好。以该条件处理果实,进行复合处理诱导果实抗病性机制的研究。
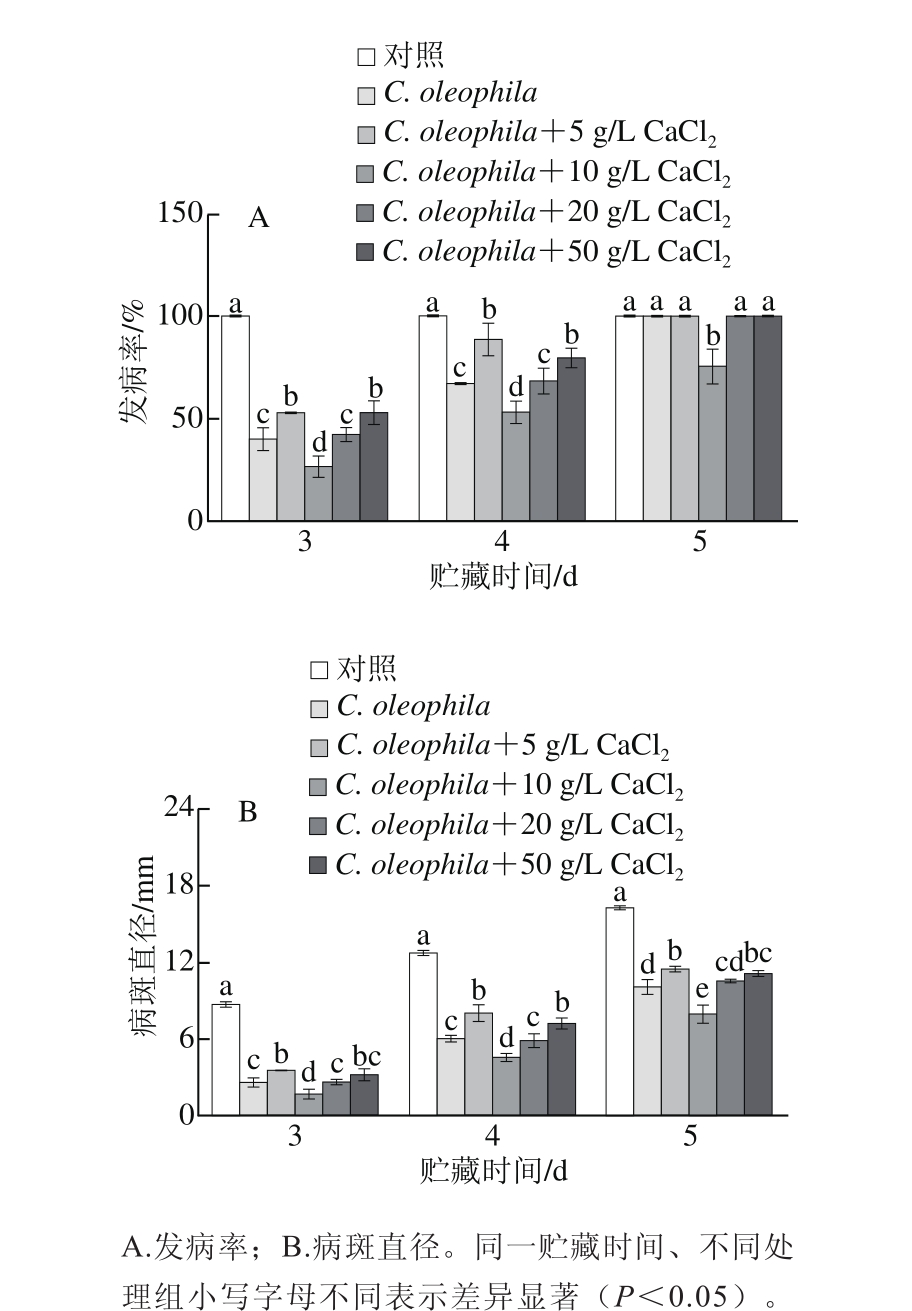
图3 CaCl2与C. oleophila复合处理对苹果果实采后青霉病的控制效果
Fig. 3 Effects of calcium chloride combined C. oleophila on disease incidence and lesion diameter in apples inoculated with P. expansum
2.2.2 CaCl2对C. oleophila在苹果果实伤口生长动态的影响
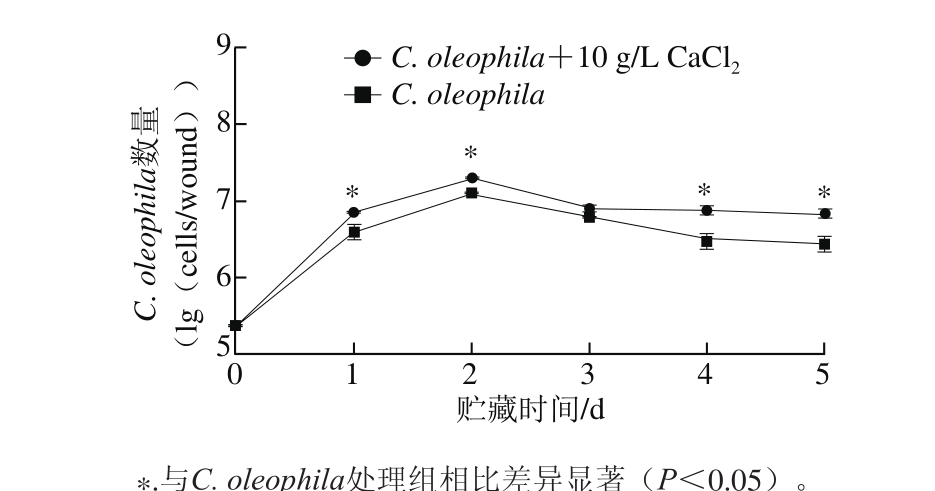
图4 CaCl2对C. oleophila在苹果果实伤口生长动态的影响
Fig. 4 Effect of calcium chloride on the growth dynamics of C. oleophila on apples
如图4所示,2 个处理组C. oleophila均能在果实伤口处迅速定植、扩增,10 g/L CaCl2处理对C. oleophila在苹果果实伤口处的生长具有促进作用,贮藏2 d时,C. oleophila+10 g/L CaCl2处理组的C. oleophila的数量比接种时增加了1.91(lg(cells/wound))。
2.3.1 CaCl2与C. oleophila复合处理对苹果果实CHI和GLU活力的影响
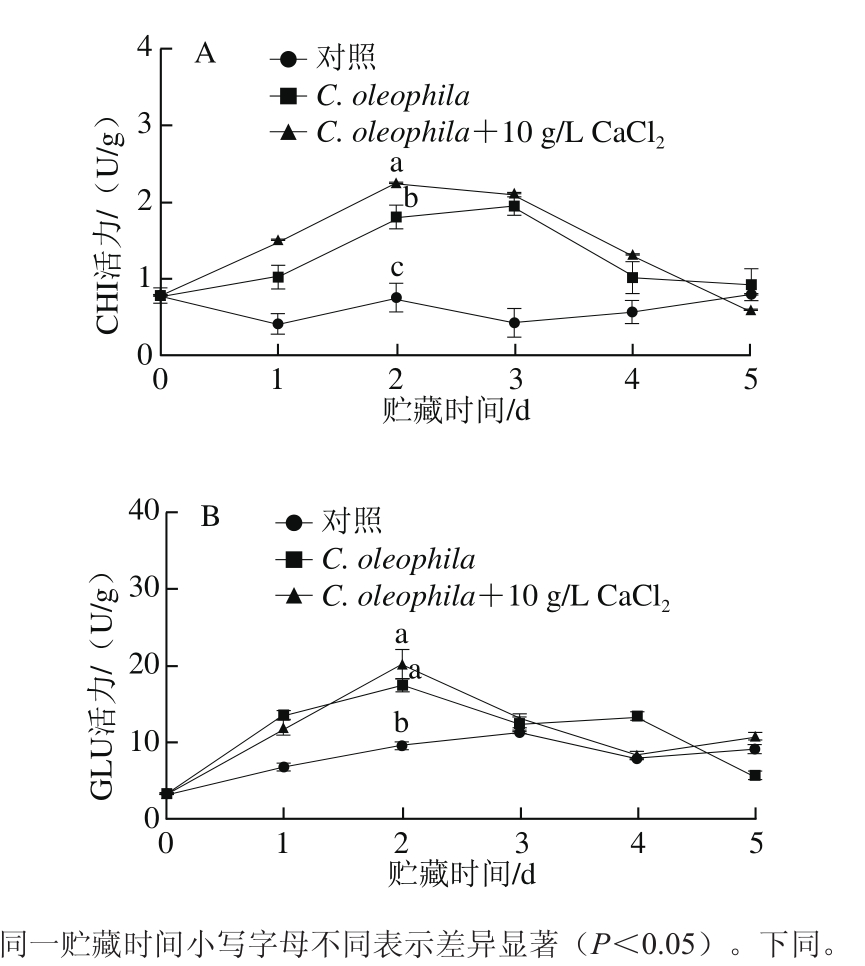
图5 CaCl2与C. oleophila复合处理对苹果果实CHI(A)和GLU(B)活力的影响
Fig. 5 Effect of calcium chloride with C. oleophila treatment on CHI (A)and GLU (B) activities in apples
CaCl2与C. oleophila复合处理对苹果果实CHI活力的影响如图5A所示。在5 d的贮藏期内,C. oleophila+10 g/L CaCl2处理组和C. oleophila单独处理组苹果果实的CHI活力大致呈先上升后下降的趋势。第2天时,C. oleophila+10 g/L CaCl2处理组苹果果实的CHI活力显著高于C. oleophila单独处理组和对照组(P<0.05)。
CaCl2与C. oleophila复合处理对苹果果实GLU活力的影响如图5B所示,在5 d的贮藏期内,C. oleophila+10 g/L CaCl2处理组苹果果实的GLU活力总体呈先上升后下降的趋势,且与对照组差异显著(P<0.05)。贮藏第2天时,C. oleophila+10 g/L CaCl2处理组苹果果实的GLU活力约为对照组的2.2 倍。
2.3.2 CaCl2与C. oleophila复合处理对苹果果实总酚、类黄酮和木质素含量的影响
CaCl2与C. oleophila复合处理对苹果果实总酚含量的影响如图6A所示。在5 d的贮藏期内,所有组苹果果实的总酚含量总体呈先上升后下降的趋势;C. oleophila+10 g/L CaCl2处理组果实总酚含量显著高于C. oleophila单独处理组和对照组(P<0.05)。贮藏第3天时,C. oleophila+10 g/L CaCl2处理组苹果果实总酚含量达到最高,约为C. oleophila处理组的1.36 倍。
CaCl2与C. oleophila复合处理对苹果果实类黄酮含量的影响如图6B所示。在5 d的贮藏期内,C. oleophila+10 g/L CaCl2处理组果实类黄酮含量显著高于C. oleophila单独处理组和对照组(P<0.05)。在贮藏第2天时,C. oleophila+10 g/L CaCl2处理组苹果果实类黄酮含量达到最大,约为C. oleophila组的1.11 倍。
CaCl2与C. oleophila复合处理对苹果果实木质素含量影响如图6C所示。在5 d的贮藏期内,C. oleophila+10 g/L CaCl2处理组果实木质素含量显著高于C. oleophila单独处理组和对照组(P<0.05),贮藏第2天时,C. oleophila+10 g/L CaCl2处理组苹果果实木质素含量达到最高。
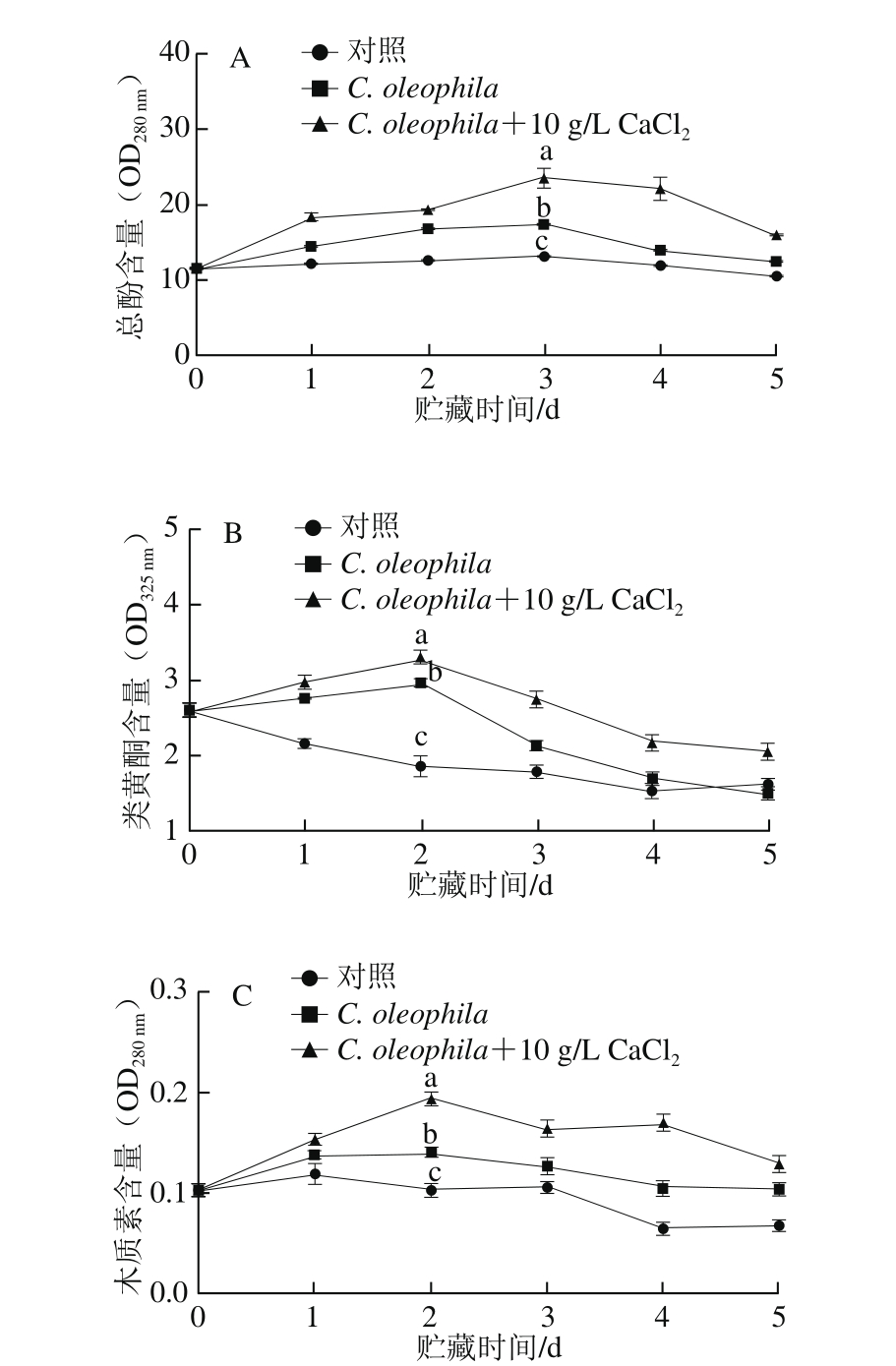
图6 CaCl2与C. oleophila处理对苹果果实总酚(A)、类黄酮(B)和木质素(C)含量的影响
Fig. 6 Effect of calcium chloride with C. oleophila treatment on the contents of total phenols (A), flavonoids (B) and lignins (C) in apples
前期研究表明,C. oleophila具备很高的商业化应用价值,但是,单一地使用拮抗酵母对果蔬采后进行生物防治的效果并不比传统的化学杀菌剂好,故而积极探索如何提高酵母的生防效力便显得尤为重要。目前,有多项研究将拮抗酵母复合其他物质(如金属离子、有机物、化学杀菌剂和其他化学或天然物质)以达到增效目的[19-24]。外源物质通过提高拮抗微生物在果实表面的生长数量,不仅可以有效地对抗病原菌的侵害,还可降低拮抗菌剂的用量,是一种更为安全有效的方式,具有很好的应用前景。
相关研究表明,果蔬在采后经过一定的物理或化学因子处理后,可产生诱导抗性,从而减少其采后病害的发生率[25-28]。CaCl2是果蔬采后处理的传统方法之一,能够对果蔬采后贮藏期有明显的延长作用[29-31]。本研究发现,C. oleophila与10 g/L CaCl2复合处理对苹果果实采后青霉病的发生抑制效果最好,10 g/L CaCl2对C. oleophila控制苹果果实采后青霉病的发生具有增效作用。
为进一步探讨CaCl2与C. oleophila复合处理对P. expansum的抑制机理,本实验测定了C. oleophila与10 g/L CaCl2复合处理对苹果相关防御酶及物质的影响。结果表明,该复合处理能有效诱导苹果相关抗性酶活力的提高。本实验选取与果蔬采后病程相关的酶及代谢产物,探究其与果蔬采后病害防治关系。病程相关蛋白家族和酚类代谢系统中的一些酶与植物体内抵抗病原微生物的侵染都有着一定的关系。酚类物质具有直接的抗菌作用[32],病原真菌的细胞壁主要成分为几丁质和葡聚糖,CHI和GLU均可通过水解病原菌细胞壁中几丁质和葡聚糖,使得病原菌原生质膜破裂从而杀死病原菌[33]。CHI和GLU在降解病原菌细胞壁时会产生协同作用,从而使抑菌活性达到最大[34]。本研究结果表明,C. oleophila与10 g/L CaCl2复合处理能显著提高苹果的CHI、GLU活力和总酚、木质素、类黄酮含量。因此,该复合处理可以通过提高防御酶活性、抗病相关蛋白活性,增加抗病物质的含量,从而诱导苹果果实采后抗病性的产生。
[1] JANISIEWICZ W J, TWORKOSKI T J, KURTZMAN C P.Biocontrol potential of Metchnikowia pulcherrima strains against blue mold of apple[J]. Phytopathology, 2001, 91(11): 1098-1108.DOI:10.1094/PHYTO.2001.91.11.1098.
[2] SPADARO D, CIAVORELLA A, DIANPENG Z, et al. Effect of culture media and pH on the biomass production and biocontrol efficacy of a Metschnikowia pulcherrima strain to be used as a biofungicide for postharvest disease control[J]. Canadian Journal of Microbiology, 2010, 56(2): 128-137. DOI:10.1139/w09-117.
[3] BENCHEQROUN S K, BAJJI M, MASSART S, et al. In vitro and in situ study of postharvest apple blue mold biocontrol by Aureobasidium pullulans: evidence for the involvement of competition for nutrients[J].Postharvest Biology and Technology, 2007, 46(2): 128-135.DOI:10.1016/j.postharvbio.2007.05.005.
[4] JANISIEWICZ W J, TWORKOSKI T J, SHARER C. Characterizing the mechanism of biological control of postharvest diseases on fruits with a simple method to study competition for nutrients[J].Phytopathology, 2000, 90(11): 1196-1200. DOI:10.1094/PHYTO.2000.90.11.1196.
[5] 范青, 田世平, 徐勇, 等. 季也蒙假丝酵母对采后桃果实软腐病的抑制效果[J]. 植物生态学报(英文版), 2000, 42(10): 1033-1038.
[6] KWASIBORSKI A, BAJJI M, RENAUT J, et al. Identification of metabolic pathways expressed by Pichia anomala Kh6 in the presence of the pathogen Botrytis cinerea on apple: new possible targets for biocontrol improvement[J]. PLoS ONE, 2014, 9(3): e91434.DOI:10.1371/journal.pone.0091434.
[7] ROMANAZZI G, SANZANI S M, BI Y, et al. Induced resistance to control postharvest decay of fruit and vegetables[J]. Postharvest Biology and Technology, 2016, 122: 82-94. DOI:10.1016/j.postharvbio.2016.08.003.
[8] SINGH S P, GAUR R. Endophytic Streptomyces spp. underscore induction of defense regulatory genes and confers resistance against Sclerotium rolfsii in chickpea[J]. Biological Control, 2017, 104: 44-56.DOI:10.1016/j.biocontrol.2016.10.011.
[9] JIN P, ZHENG C, HUANG Y P, et al. Hot air treatment activates defense responses and induces resistance against Botrytis cinerea in strawberry fruit[J]. Journal of Integrative Agriculture, 2016, 15(11):2658-2665. DOI:10.1016/S2095-3119(16)61387-4.
[10] XU X, QIN G, TIAN S. Effect of microbial biocontrol agents on alleviating oxidative damage of peach fruit subjected to fungal pathogen[J]. International Journal of Food Microbiology, 2008,126(1/2): 153-158. DOI:10.1016/j.ijfoodmicro.2008.05.019.
[11] 邵兴锋, 屠康, 静玮, 等. 热处理对红富士苹果贮藏期间青霉病的抑制效果[J]. 园艺学报, 2007, 34(3): 743-746. DOI:10.16420/j.issn.0513-353x.2007.03.037.
[12] BOLLER T, GEHRI A, MAUCH F, et al. Chitinase in bean leaves: induction by ethylene, purification, properties, and possible function[J]. Planta, 1983, 157(1): 22-31.
[13] ABELES F B, FORRENCE L E. Temporal and hormonal control of β-1,3-glucanase in Phaseolus vulgaris L.[J]. Plant Physiology, 1970,45(4): 395-400.
[14] LI Hua, SUO Jiangtao, HAN Ye, et al. The effect of 1-methylcyclopropene, methyl jasmonate and methyl salicylate on lignin accumulation and gene expression in postharvest ‘Xuxiang’kiwifruit during cold storage[J]. Postharvest Biology & Technology,2017, 124: 107-118. DOI:10.1016/j.postharvbio.2016.10.003.
[15] DENG Lili, ZENG Kaifang, ZHOU Yahan, et al. Effects of postharvest oligochitosan treatment on anthracnose disease in citrus (Citrus sinensis L. Osbeck) fruit[J]. European Food Research and Technology, 2015,240(4): 795-804. DOI:10.1007/s00217-014-2385-7.
[16] DIXON R A, PALVA N L. Stress-induced phenylpropanoid metabolism[J]. The Plant Cell, 1995, 7(7): 1085-1097.
[17] HUCKELHOVEN R, FODOR J, PREIS C, et al. Hypersensitive cell death and papilla formation in barley attacked by the powdery mildew fungus are associated with hydrogen peroxide but not with salicylic acid accumulation[J]. Plant Physiology, 1999, 119(4): 1251-1260.
[18] YAO H J, TIAN S P. Effects of pre- and post-harvest application of salicylic acid or methyl jasmonate on inducing disease resistance of sweet cherry fruit in storage[J]. Postharvest Biology & Technology,2005, 35(3): 253-262. DOI:10.1016/j.postharvbio.2004.09.001.
[19] VERO S, MONDINO P, BURGUEÑO J, et al. Characterization of biocontrol activity of two yeast strains from Uruguay against blue mold of apple[J]. Postharvest Biology & Technology, 2002, 26(1): 91-98. DOI:10.1016/S0925-5214(01)00199-5.
[20] DROBY S, WISNIEWSKI M, MACARISIN D, et al. Twenty years of postharvest biocontrol research: is it time for a new paradigm?[J].Postharvest Biology and Technology, 2009, 52(2): 137-145.DOI:10.1016/j.postharvbio.2008.11.009.
[21] 余挺. 提高罗伦隐球酵母拮抗效力的途径及其机理的研究[D].杭州: 浙江大学, 2007: 20.
[22] QIN G Z, TIAN S P. Enhancement of biocontrol activity of Cryptococcus laurentii by silicon and the possible mechanisms involved[J]. Phytopathology, 2008, 95(1): 69-75. DOI:10.1094/PHYTO-95-0069.
[23] SUI Y, LIU J. Effect of glucose on thermotolerance and biocontrol efficacy of the antagonistic yeast Pichia guilliermondii[J]. Biological Control, 2014, 74(3): 59-64. DOI:10.1016/j.biocontrol.2014.04.003.
[24] 冯吉睿, 周雅涵, 曾凯芳. 膜醭毕赤酵母结合杀菌剂对柑橘果实采后病害的控制效果[J]. 食品科学, 2015, 36(12): 249-254.
[25] 王艳颖, 胡文忠, 庞坤, 等. 机械损伤对富士苹果生理生化变化的影响[J]. 食品与发酵工业, 2007, 33(7): 58-62. DOI:10.13995/j.cnki.11-1802/ts.2007.07.018.
[26] 吕敏, 苏建坤, 白和盛, 等. 桃蚜取食和机械损伤对番茄和辣椒PAL、LOX和PPO活性的诱导作用[J]. 江苏农业学报, 2016, 32(6):1273-1279.
[27] 曾凯芳, 姜微波. 水杨酸处理对采后绿熟芒果炭疽病抗病性的诱导[J].中国农业大学学报, 2005, 10(2): 36-40.
[28] 林丽, 田世平, 秦国政, 等. 两种拮抗酵母菌对桃果实贮藏期间主要病害的防治效果[J]. 中国农业科学, 2003, 36(12): 1535-1539.
[29] 陈敏, 高云慨, 宋海超, 等. 氯化钙结合季也蒙毕赤酵母(Meyerozyma guilliermondii)对抑制芒果采后炭疽病效果的影响[J]. 食品科学,2016, 37(2): 204-209.
[30] 陈莉, 郝浩永, 程朝霞, 等. 采后氯化钙处理对番茄生理的影响[J].长江蔬菜, 2009(8): 27-29.
[31] 李玉萍. 氯化钙对番石榴果实采后品质的影响[J]. 世界热带农业信息, 2003(11): 23-24.
[32] NICHOLSON R L N, HAMMERSCHMIDT R. Phenolic compounds and their role in disease resistance[J]. Annual Review of Phytopathology, 1992, 30: 369-389.
[33] SCHLUMBAUM A, MAUCH F, VÖGELI U, et al. Plant chitinases are potent inhibitors of fungal growth[J]. Nature, 1986, 324: 365-367.
[34] VAN LOON L C. Induced resistance in plants and the role of pathogenesis-related proteins[J]. European Journal of Plant Pathology,1997, 103(9): 753-765.
CaCl2 Enhances the Biocontrol Efficacy of Candida oleophila against Inoculated Penicillium expansum on Postharvest Apples
蔡孟轩, 邓丽莉, 姚世响, 等. CaCl2增强橄榄假丝酵母对苹果果实采后青霉病的生防效力[J]. 食品科学, 2019, 40(11):220-226. DOI:10.7506/spkx1002-6630-20180530-427. http://www.spkx.net.cn
CAI Mengxuan, DENG Lili, YAO Shixiang, et al. CaCl2 enhances the biocontrol efficacy of Candida oleophila against inoculated Penicillium expansum on postharvest apples[J]. Food Science, 2019, 40(11): 220-226. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-20180530-427. http://www.spkx.net.cn