 表示,指标比较采用最小显著差异法,取95%置信度(P<0.05)。采用Origin 7.5软件作图。
表示,指标比较采用最小显著差异法,取95%置信度(P<0.05)。采用Origin 7.5软件作图。米糠是稻谷加工过程中最重要的一类副产物,虽然只占稻谷质量的6%~8%,但却集中了稻谷中60%~70%的营养素,包括丰富的脂质、蛋白质等[1]。然而,米糠极易酸败,导致米糠及其制品营养品质下降[2]。长期以来,关于米糠酸败影响米糠制品营养品质的研究主要围绕米糠油展开,而忽略了米糠蛋白营养品质的变化[3]。米糠蛋白在脱脂米糠中质量占比达15%~20%,米糠蛋白致敏性低、氨基酸组成合理、生物效价高,是一种极具开发潜力的新型植物蛋白,特别适合开发婴幼儿配方食品[4]。近期有研究表明,米糠氧化酸败产物可导致米糠蛋白氧化,使得米糠蛋白结构特征和功能性质发生改变[5]。随着米糠酸败程度的增加,米糠蛋白体外胃蛋白酶消化产物的抗氧化性也会发生显著变化[6]。蛋白质消化产物往往直接被机体吸收与利用,蛋白质消化产物抗氧化性的改变势必影响机体氧化还原状态,从而可能诱发机体氧化应激[7-8]。为全面评价和控制米糠酸败对米糠蛋白营养品质的影响,研究米糠酸败影响米糠蛋白消化产物抗氧化性的机理尤为重要,而蛋白质消化产物的抗氧化性与其结构特征(分子质量大小、表面疏水性等)密切相关[9-11]。因此,有必要对米糠酸败过程中米糠蛋白消化产物结构特征的变化进行深入分析。本研究以新鲜米糠为原料,在室温下贮藏一定时间得到不同酸败程度的米糠,经过稳定化和脱脂处理后制备米糠蛋白,研究米糠酸败对米糠蛋白体外胃蛋白酶消化产物结构特征的影响,以期为开发米糠蛋白及相关健康食品提供参考。
新鲜米糠(酸值3 mg/g) 湖南粮食集团有限责任公司;胃蛋白酶(800 U/mg) 合肥Biosharp公司;十二烷基硫酸钠(sodium dodecyl sulfate,SDS)、丙烯酰胺、N,N’-甲叉双丙烯酰胺、过硫酸铵 美国Sigma公司;标准分子质量蛋白 北京全式金生物技术有限公司;三羟甲基氨基甲烷(Tris)、β-巯基乙醇、乙酸等(均为分析纯) 国药集团化学试剂有限公司。
RXZ-128A型人工气候箱 宁波市科技园区新江南仪器有限公司;FMHE36-24型双螺杆挤压机 湖南富马科食品工程技术有限公司;Sorvall LYNX6000型冷冻离心机 美国Thermo Fisher公司;FD5-4T型冷冻干燥机金西盟(北京)仪器有限公司;DSI Z-300A型水浴恒温振荡器 苏州培英实验设备有限公司;Nano ZS型纳米粒度分析仪 英国Malvern公司;F-4600型荧光分光光度计 日本日立公司;LC-20A型高效液相色谱仪日本岛津公司;SE260型电泳仪 美国GE公司。
1.3.1 新鲜米糠贮藏不同时间制备不同酸败程度的米糠
参考何莉媛等[6]的方法,以新鲜米糠为原料粉碎过40 目筛,在室温下分别贮藏0、1、3、5、10 d得到不同酸败程度的米糠,采用双螺杆挤压机进行稳定化处理。稳定化条件为:进料量15 kg/h,控制水分质量分数16%,双螺杆挤压机2~6区温度依次为70、120、120、70 ℃和60 ℃。然后,将稳定化处理的米糠与正己烷按料液比1∶4(m/V)混合脱脂,收集米糠毛油,所得米糠毛油酸值分别为3、16、26、35、40 mg/g。最后,将滤饼置于通风橱中风干后得到脱脂米糠,将脱脂米糠置于4 ℃冰箱保存备用。
1.3.2 米糠蛋白的制备
参考吴伟等[5]的方法,将脱脂米糠与去离子水按1∶10(m/V)混合,用2 mol/L NaOH溶液调pH值至9.0,40 ℃下搅拌反应4 h。将悬浮液在4℃、8 000 r/min离心15 min,取上清液用2 mol/L HCl溶液调pH值至4.0,静置30 min后在4 ℃、8 000 r/min离心15 min得到米糠蛋白沉淀。水洗沉淀3 次,再将其分散于去离子水中,用2 mol/L NaOH溶液调pH值至7.0,冷冻干燥得到米糠蛋白。
1.3.3 米糠蛋白体外胃蛋白酶消化样品的制备
参考Chen Nannan等[12]的方法,分别将不同贮藏时间米糠制备的米糠蛋白用去离子水配制成质量分数2%的米糠蛋白悬浮液,将悬浮液置于37 ℃水浴30 min,并用2 mol/L HCl溶液调pH值至2.0。然后,分别加入4 U/mg胃蛋白酶,在37 ℃进行体外消化反应。消化0、5、10、20、30、40、60 min和90 min后取样,用2 mol/L NaOH溶液将消化不同时间的米糠蛋白悬浮液调pH值至7.0,终止消化后对所得样液进行冷冻干燥,于4 ℃冰箱保存备用。
1.3.4 米糠蛋白消化产物亚基结构的测定
参考吴伟等[5]的方法,采用SDS-聚丙烯酰胺凝胶电泳法。样品质量浓度为3 mg/mL,上样量为12 μL,分离胶和浓缩胶质量分数分别为12.5%和4%,电极缓冲液含0.05 mol/L Tris、0.384 mol/L甘氨酸、0.1% SDS(pH 8.3),样品溶解液含SDS 2 g/100 mL、β-巯基乙醇5 g/100 mL、甘油10 g/100 mL、溴酚蓝0.02 g/100 mL、0.01 mol/L pH 8.0 Tris-HCl缓冲液。固定液为454 mL体积分数50%甲醇溶液与46 mL冰乙酸混合液,染色液为考马斯亮蓝R-250,脱色液为体积分数95%乙酸溶液。电泳前期的电流为10 mA,待样品进入分离胶改为25 mA。
1.3.5 米糠蛋白消化产物分子质量分布的测定
参考Chen Nannan等[12]的方法,用高效液相色谱仪测定米糠蛋白消化产物的分子质量分布。将米糠蛋白消化产物(1.0 mg/mL)用磁力搅拌器搅拌2 h,4 ℃、10 000 r/min离心20 min,取上清液用0.45 μm醋酸纤维素膜过滤备用。色谱柱:TSKgel SW G2000 SWXL(300 mm×7.8 mm,5 μm);检测器:Waters 996光电二极管阵列检测器;流动相:0.1 mol/L磷酸盐缓冲液(pH 6.8,含0.1 mol/L Na2SO4);紫外检测波长:214 nm;流速:0.5 mL/min;柱温:25 ℃。蛋白质标准曲线所用5 种标准品:细胞色素C(12 384 u)、抑肽酶(6 511 u)、杆菌肽(1 450 u)、氧化型谷胱甘肽(651 u)、Gly-Gly-Tyr-Arg(451 u)、Gly-Gly-Gly(189 u),蛋白质标准曲线为lg Y=-0.237 1X+7.157 7(R2=0.985 6),其中Y为蛋白质相对分子质量,X为保留时间/min。
1.3.6 米糠蛋白消化产物粒径分布的测定
参考吴伟等[5]的方法,采用纳米粒度分析仪测定米糠蛋白消化产物的粒径分布,样品质量浓度为0.25 mg/mL。
1.3.7 米糠蛋白消化产物荧光峰位的测定
参考Wu Wei等[13]的方法,用考马斯亮蓝法测定米糠蛋白消化产物的蛋白质量浓度,将其稀释至0.1 mg/mL,并用F-4600型荧光分光光度计在激发波长280 nm处得到300~500 nm波长之间的发射光谱。
1.3.8 米糠蛋白消化产物表面疏水性的测定
参考Huang Youru等[14]的方法,采用ANS荧光探针法测定米糠蛋白消化产物的表面疏水性。稀释蛋白质量浓度梯度为0.005、0.01、0.02、0.05、0.08 mg/mL和0.10 mg/mL;激发波长和发射波长分别为384 nm和471 nm,灵敏度为6。以荧光强度对样品质量浓度作图,外推至蛋白质质量浓度为0,曲线初始阶段的斜率即为米糠蛋白消化产物的表面疏水性指数。
所有实验平行测定3 次。数据采用Microsoft Excel 2003和Origin 7.5软件进行处理,结果以表示,指标比较采用最小显著差异法,取95%置信度(P<0.05)。采用Origin 7.5软件作图。
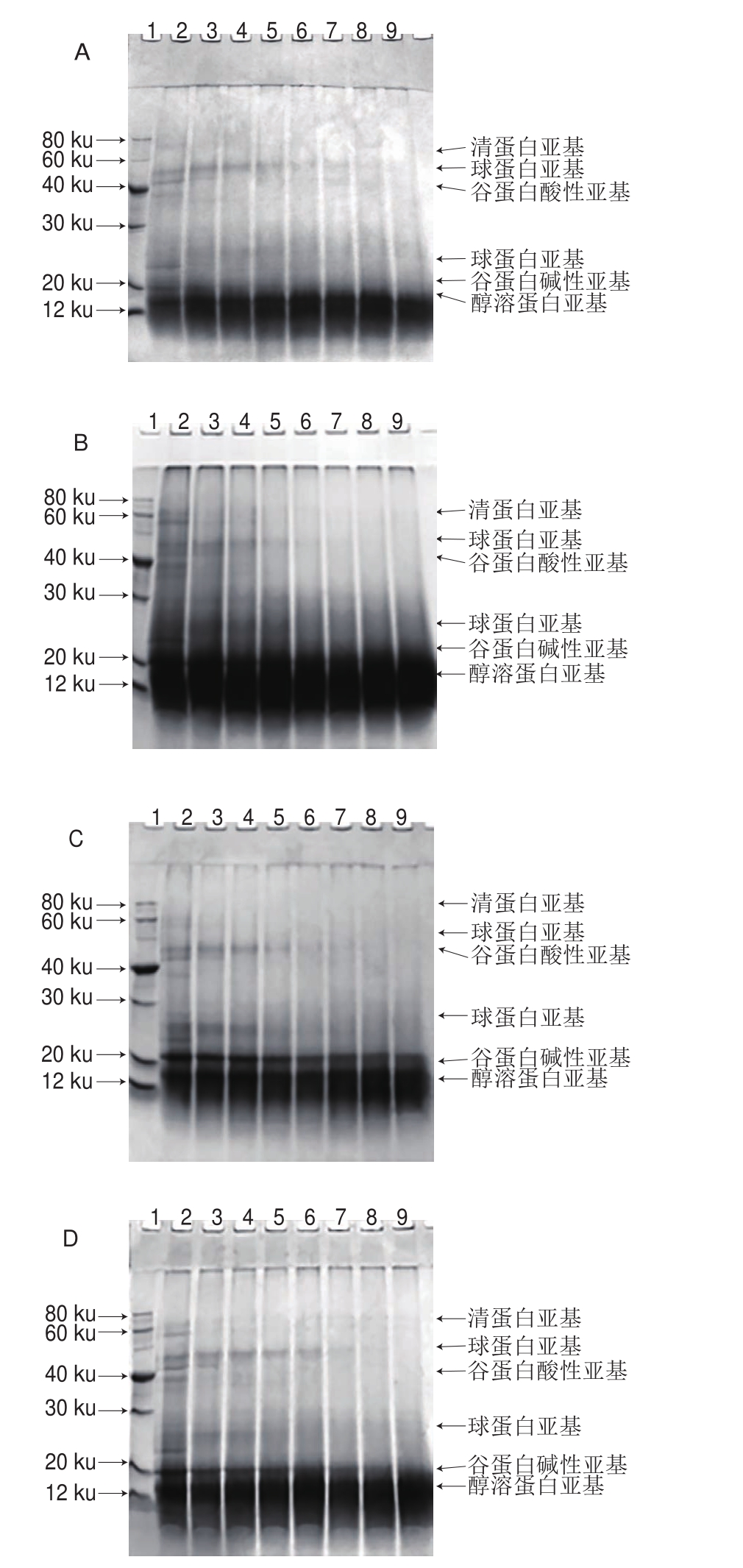

图1 米糠酸败对米糠蛋白体外胃蛋白酶消化产物亚基结构的影响
Fig. 1 Effect of rice bran rancidity on subunit structures of in vitro pepsin digest of rice bran protein
米糠蛋白是一种混合蛋白,包含清蛋白、球蛋白、谷蛋白和醇溶蛋白,其亚基结构十分复杂,米糠清蛋白主要以高分子质量亚基为主,米糠球蛋白既有高分子质量亚基也有低分子质量亚基,米糠谷蛋白和米糠醇溶蛋白则以低分子质量亚基为主[15-16]。在体外胃蛋白酶消化过程中,随着胃蛋白酶消化时间延长,高分子质量亚基和低分子质量亚基均发生了不同程度的降解。从图1A可以看出,米糠清蛋白亚基(60~80 ku)和米糠谷蛋白酸性亚基(37 ku左右)最先被胃蛋白酶完全消化降解,酶解10 min左右,二者对应的亚基条带均已消失;随后米糠谷蛋白碱性亚基(20 ku左右)和米糠球蛋白亚基(20~30、40~60 ku)也逐渐被胃蛋白酶消化降解;而米糠醇溶蛋白亚基(7~14 ku)对应的电泳条带则先变宽后变窄。这表明,米糠谷蛋白酸性亚基比碱性亚基容易被胃蛋白酶消化,亲水性亚基比疏水性亚基容易被胃蛋白酶消化。江连洲等[17]关于体外胃蛋白酶消化大豆蛋白的研究也发现,大豆球蛋白中酸性亚基比碱性亚基容易被胃蛋白酶降解,β-伴大豆球蛋白中带有较强亲水性“肽链”的α-亚基,比具有疏水性“核心”的β-亚基容易被胃蛋白酶降解。体外胃蛋白酶消化过程中,随着米糠酸败程度加剧,米糠清蛋白亚基、谷蛋白酸性亚基和球蛋白亚基完全降解的时间先推前后延迟,米糠贮藏3 d制备的米糠蛋白最容易被消化,消化时间30 min时,这些亚基对应的电泳条带基本都已消失(图1C)。而米糠酸败过程中,米糠谷蛋白碱性亚基和醇溶蛋白亚基则总体上表现为越来越难被胃蛋白酶消化,米糠贮藏10 d制备的米糠蛋白被消化90 min时,这两种亚基对应的电泳条带依然存在(图1E)。关于米糠酸败对米糠蛋白体外胃蛋白消化进程影响的研究也表明,米糠贮藏3 d制备的米糠蛋白初始消化速率最大,而米糠贮藏10 d制备的米糠蛋白初始消化速率最小[6]。这可能是由于米糠贮藏3 d时酸败程度较低,产生的米糠氧化酸败产物可诱导蛋白质分子部分去折叠,空间结构柔性增加,暴露出更多的胃蛋白酶酶解位点,使蛋白质容易被消化[18]。随着米糠贮藏时间的延长,米糠氧化酸败程度快速增加,产生大量的脂质自由基和活性脂质氧化产物[19]。关于蛋白质氧化的大量研究已证实,过量的过氧自由基[20]、丙二醛[21]等均会诱导蛋白质氧化形成共价交联聚集体。图1E也显示,米糠贮藏10 d制备的米糠蛋白形成了大量共价交联聚集体,随着消化时间延长,蛋白质聚集体被降解成与天然亚基分子质量大小类似的亚基,但是这些亚基的空间结构在去折叠-聚集-酶解的过程中发生了改变,可能使米糠蛋白氧化初期暴露的胃蛋白酶酶解位点被掩蔽,从而难以被胃蛋白酶消化降解。
体外胃蛋白酶消化过程中,蛋白质分子通常先降解成䏡和胨,再水解为小分子肽,其分子质量变化是一个动态过程[22]。如表1所示,随着胃蛋白酶消化时间的延长,米糠蛋白消化产物中大于10 ku的组分均急剧减少,3~10 ku的组分均先增加后减少,而小于3 ku的组分则均持续增加。蛋白酶一般具有较广泛的作用位点,然而胃蛋白酶是专一性较强的蛋白酶,通常被认为不易将蛋白质水解成小分子肽[23]。Chen Nannan等[24]采用胃蛋白酶水解天然大豆蛋白1 h得到的消化产物中小于3 ku的蛋白肽仅为19.64%,远低于同等条件下本实验制备的米糠蛋白消化产物(44.05%~50.04%)。这表明,相比于大豆蛋白,米糠蛋白易于被胃蛋白酶消化降解成小分子肽,并且有文献报道,这些小分子肽是米糠蛋白消化产物具有抗氧化活性的关键组分[9-10]。
表1 米糠酸败对米糠蛋白体外胃蛋白酶消化产物分子量分布的影响
Table 1 Effect of rice bran rancidity on molecular mass distribution of in vitro pepsin digest of rice bran protein
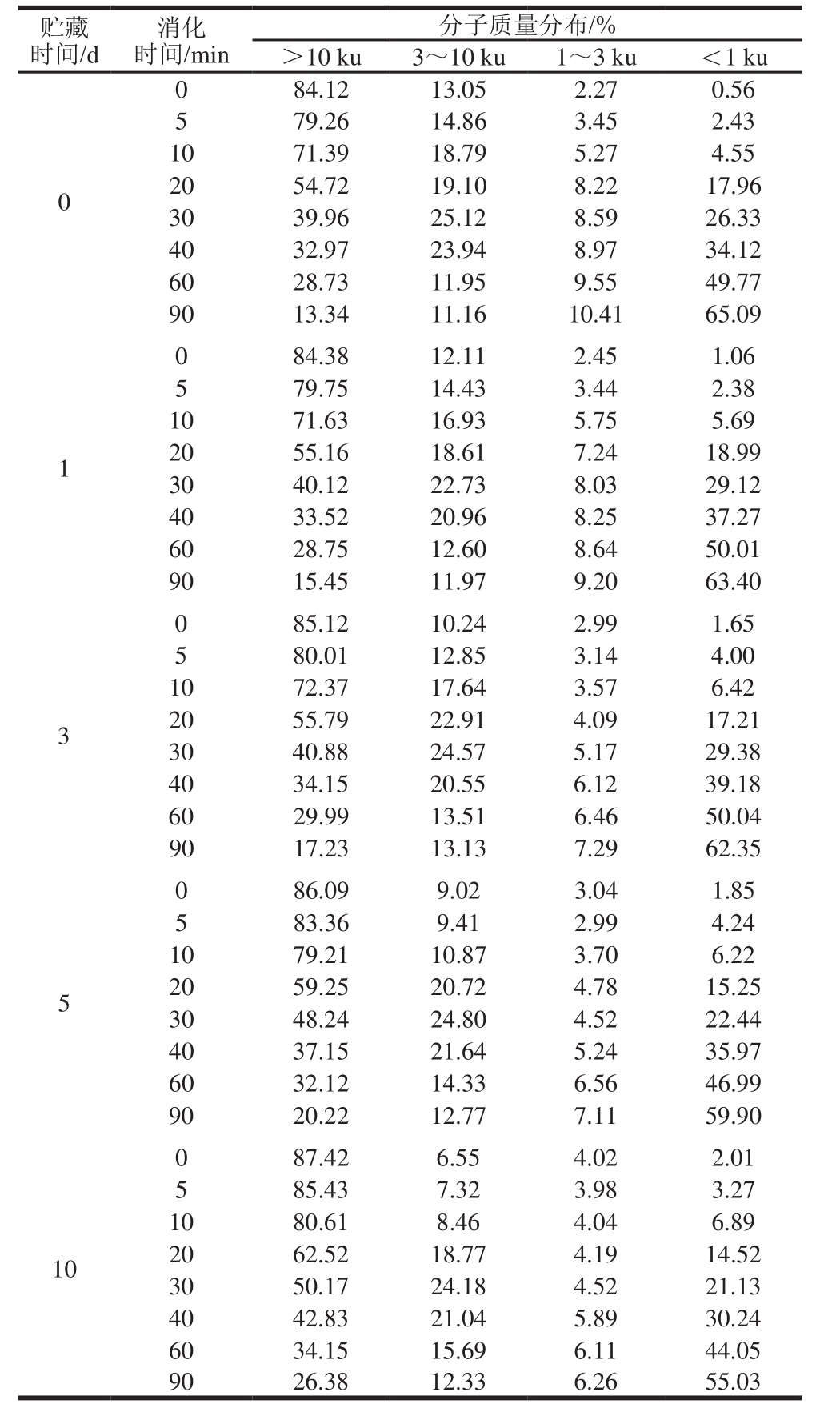
贮藏时间/d消化时间/min分子质量分布/%>10 ku 3~10 ku 1~3 ku <1 ku 0 0 84.12 13.05 2.27 0.56 5 79.26 14.86 3.45 2.43 10 71.39 18.79 5.27 4.55 20 54.72 19.10 8.22 17.96 30 39.96 25.12 8.59 26.33 40 32.97 23.94 8.97 34.12 60 28.73 11.95 9.55 49.77 90 13.34 11.16 10.41 65.09 1 0 84.38 12.11 2.45 1.06 5 79.75 14.43 3.44 2.38 10 71.63 16.93 5.75 5.69 20 55.16 18.61 7.24 18.99 30 40.12 22.73 8.03 29.12 40 33.52 20.96 8.25 37.27 60 28.75 12.60 8.64 50.01 90 15.45 11.97 9.20 63.40 3 0 85.12 10.24 2.99 1.65 5 80.01 12.85 3.14 4.00 10 72.37 17.64 3.57 6.42 20 55.79 22.91 4.09 17.21 30 40.88 24.57 5.17 29.38 40 34.15 20.55 6.12 39.18 60 29.99 13.51 6.46 50.04 90 17.23 13.13 7.29 62.35 5 0 86.09 9.02 3.04 1.85 5 83.36 9.41 2.99 4.24 10 79.21 10.87 3.70 6.22 20 59.25 20.72 4.78 15.25 30 48.24 24.80 4.52 22.44 40 37.15 21.64 5.24 35.97 60 32.12 14.33 6.56 46.99 90 20.22 12.77 7.11 59.90 10 0 87.42 6.55 4.02 2.01 5 85.43 7.32 3.98 3.27 10 80.61 8.46 4.04 6.89 20 62.52 18.77 4.19 14.52 30 50.17 24.18 4.52 21.13 40 42.83 21.04 5.89 30.24 60 34.15 15.69 6.11 44.05 90 26.38 12.33 6.26 55.03
从表1还可以看出,未水解的米糠蛋白分子质量主要集中在10 ku以上,占比达84%以上。随着米糠贮藏时间的延长,未水解的米糠蛋白大于10 ku和小于3 ku的组分均略微增加,而3~10 ku的组分则有所减少。结合前面的SDS-聚丙烯酰胺凝胶电泳图分析,大于10 ku组分增加的原因应该是米糠脂质酸败氧化产物诱导蛋白质氧化形成了可溶性的聚集体,并且吴伟等[5]关于米糠酸败对米糠蛋白结构影响的研究已证实,形成的主要是大于1 000 ku的高分子质量聚集体。小于3 ku组分也增加可能是因为部分米糠脂质酸败氧化产物如过氧自由基等,不仅可诱导蛋白质氧化聚集,同时还可切断蛋白质主肽链,形成分子质量较小的肽类物质[25]。有关大豆蛋白[26]和猪肌原纤维蛋白[27]的氧化研究也发现,较高浓度的脂质过氧化产物可诱导蛋白质组分部分降解。酶解90 min时得到的米糠蛋白消化产物的分子质量分布则显示,分子质量主要集中在1 ku以下,占比达55%以上。随着米糠贮藏时间的延长,酶解90 min时得到的米糠蛋白消化产物中大于10 ku的组分占比增加,而小于3 ku的多肽占比变小,这同样表明,米糠酸败过程中米糠蛋白形成的氧化聚集体难以被胃蛋白酶降解成小分子肽。
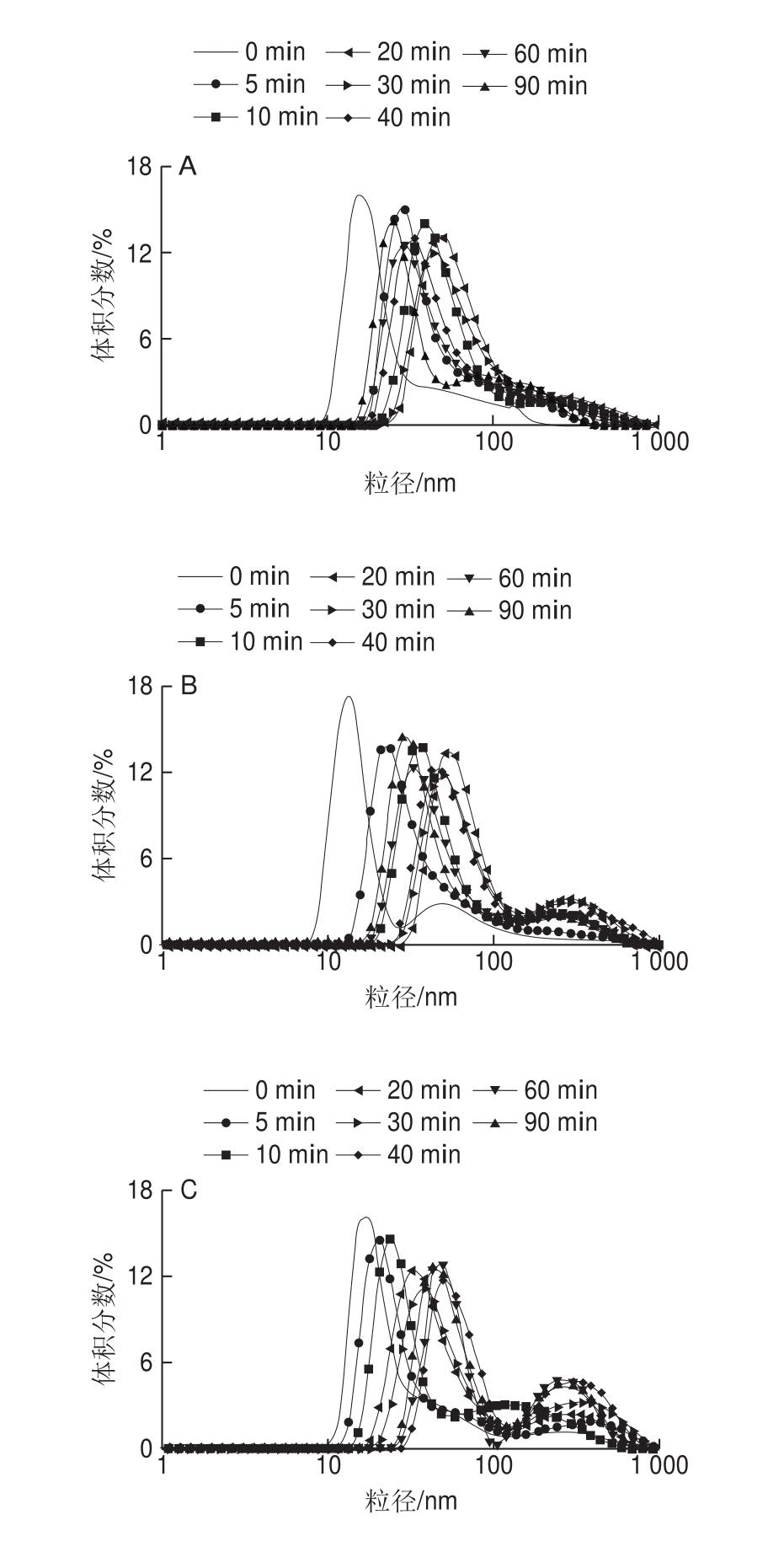

图2 米糠酸败对米糠蛋白体外胃蛋白酶消化产物粒径分布的影响
Fig. 2 Effect of rice bran rancidity on particle size distribution of in vitro pepsin digest o rice bran protein
米糠蛋白消化产物的粒径分布如图2所示,随着胃蛋白酶消化时间延长,不同酸败程度米糠制备的米糠蛋白消化产物的粒径分布均先往大尺寸方向偏移,再往小尺寸方向偏移,在消化30 min时粒径分布曲线位于最大尺寸处。平均粒径结果也显示,随着消化时间延长,5 组贮藏不同时间米糠制备的米糠蛋白消化产物平均粒径均先增加后减小(图3)。Cui Chun等[28]关于不同pH值条件下胃蛋白酶消化大豆蛋白的研究同样发现,随着胃蛋白酶消化时间的延长,大豆蛋白消化产物的平均粒径先增大后减小,并认为消化前期粒径增大的原因是胃蛋白酶水解诱导大豆球蛋白去折叠形成了大的可溶性聚集体,消化后期粒径减小的原因是聚集体逐渐降解为低分子质量肽。结合前面分子质量分布的结果可以推断,随着消化时间进一步延长,米糠蛋白消化产物粒径减小的原因也是由于形成了较多的低分子质量肽。
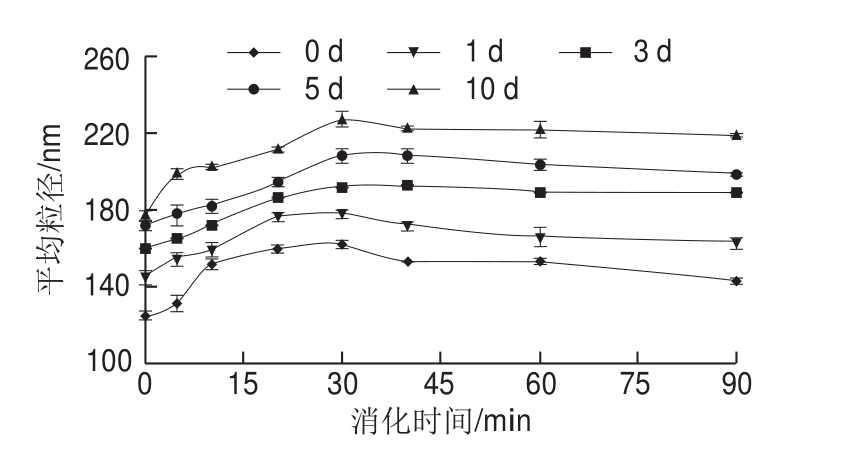
图3 米糠酸败对米糠蛋白体外胃蛋白酶消化产物平均粒径的影响
Fig. 3 Effect of rice bran rancidity on average particle size of in vitro pepsin digest of rice bran protein
体外胃蛋白酶消化过程中,随着米糠贮藏时间的延长,米糠蛋白消化产物的粒径分布曲线由单峰分布(图2A)逐渐变成双峰分布(图2B~E),且在100~1 000 nm之间峰所占体积分数逐渐增加。平均粒径结果显示,当消化时间相同时,随着米糠贮藏时间延长,米糠蛋白消化产物的平均粒径越大(图3)。吴伟等[5]关于米糠酸败对米糠蛋白结构影响的研究已证实,随着米糠贮藏时间的延长,米糠蛋白粒径增加是由于可溶性聚集体的形成。米糠蛋白消化产物的粒径分布结果进一步表明,米糠酸败可导致米糠蛋白形成氧化聚集体,且这类氧化聚集体难以被胃蛋白酶消化分解。
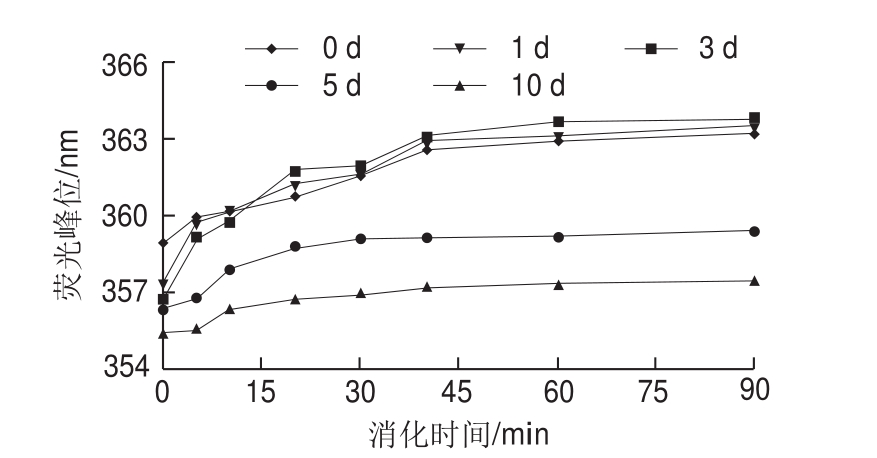
图4 米糠酸败对米糠蛋白体外胃蛋白酶消化产物内源荧光光谱峰位的影响
Fig. 4 Effect of rice bran rancidity on maximum emission wavelength in intrinsic fluorescence spectra of in vitro pepsin digest of rice bran protein
蛋白质内源荧光光谱主要反映蛋白质芳香族氨基酸残基及其微环境的变化[29-30]。米糠蛋白消化产物的荧光峰位与消化时间关系曲线如图4所示,随着体外胃蛋白酶消化时间延长,5 组贮藏不同时间米糠制备的米糠蛋白消化产物荧光峰位均发生红移。这可能是由于胃蛋白酶具有一定的氨基酸序列选择特异性,优先断裂由芳香族氨基酸(苯丙氨酸、酪氨酸和色氨酸)或亮氨酸形成的肽键,随着消化时间延长,大量芳香族氨基酸从蛋白质内部的疏水性中心转移到外部的亲水性环境中[23]。Cui Chun等[28]关于不同pH值条件下胃蛋白酶消化大豆蛋白的研究也发现,随着消化时间延长,大豆蛋白荧光峰位发生红移,同样认为是由于芳香族氨基酸残基的微环境由疏水环境转变成了亲水环境。随着米糠贮藏时间的延长,米糠蛋白消化产物的红移幅度先增大后减小,在第3天时红移幅度最大。何莉媛等[6]在研究米糠酸败对米糠蛋白体外胃蛋白酶消化性质的影响时发现,随着米糠贮藏时间延长,米糠蛋白消化率和消化速率均呈现先上升后下降的趋势,也在米糠贮藏3 d 时达到最大值。由此推测,当米糠酸败程度较低时,米糠蛋白发生适度氧化,有利于蛋白质分子在胃蛋白酶消化过程中结构展开,使更多内部的芳香族氨基酸残基暴露于亲水环境中,使得荧光峰位红移幅度增加;当米糠酸败程度较高时,米糠蛋白过度氧化形成的共价交联聚集体使得胃蛋白酶作用位点减少,不利于内部的芳香族氨基酸残基被水解,使得荧光峰位红移幅度减小。
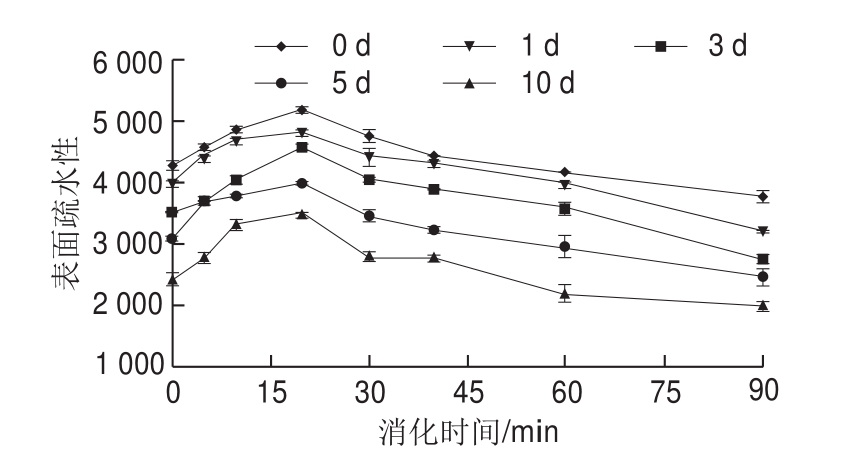
图5 米糠酸败对米糠蛋白体外胃蛋白酶消化产物表面疏水性的影响
Fig. 5 Effect of rice bran rancidity on surface hydrophobicity of in vitro pepsin digest of rice bran protein
表面疏水性是指在极性溶液中蛋白质分子表面疏水性氨基酸的相对含量,被作为评价蛋白质结构变化的重要参数[18]。米糠蛋白消化产物的表面疏水性如图5所示,随着体外胃蛋白酶消化时间延长,5 组贮藏不同时间米糠制备的米糠蛋白消化产物的表面疏水性均先上升后下降,在消化20 min左右达到最大值。Cui Chun等[28]关于大豆蛋白胃蛋白酶消化的研究也发现,随着消化时间延长,消化产物的表面疏水性先上升后下降。Radha等[31]研究发现,采用蛋白酶水解20 min得到的大豆蛋白酶解产物的表面疏水性相比于未改性的大豆蛋白显著增加,归因于酶解促使掩埋于大豆球蛋白分子内部的疏水性基团暴露到外部亲水性环境中。Bamdad等[32]则发现,当水解时间超过30 min时,大麦醇溶蛋白消化产物的表面疏水性逐渐下降,归因于醇溶蛋白分子内部掩埋了更多亲水性基团,酶解诱导其内部的亲水性基团暴露于外部。结合前面米糠蛋白消化产物的电泳图分析,消化过程中表面疏水性先升后降的原因是:消化初期,胃蛋白酶首先降解米糠清蛋白、球蛋白等亲水性亚基,使内部的疏水性基团暴露;消化后期,米糠醇溶蛋白也逐渐被水解,诱导其内部的亲水性基团暴露。此外,暴露的疏水性基团通过疏水相互作用聚集也会导致表面疏水性下降[28]。随着米糠贮藏时间越长,米糠蛋白及其消化产物的表面疏水性均越来越小。吴伟等[5]认为米糠酸败导致米糠蛋白表面疏水性下降的主要原因,一是酸败过程产生的脂质氧化产物与米糠蛋白的疏水基团发生了反应,二是进一步通过疏水相互作用形成了氧化聚集体。这可能也是造成米糠蛋白胃蛋白酶消化产物的表面疏水性随着米糠酸败程度增加不断下降的原因。
将新鲜米糠贮藏一定时间得到不同酸败程度的米糠,随后脱脂制备米糠蛋白,研究米糠酸败过程中米糠蛋白体外胃蛋白酶消化产物结构特征的变化。电泳分析显示,在体外胃蛋白酶消化过程中,新鲜米糠的清蛋白亚基和谷蛋白酸性亚基最先被胃蛋白酶完全消化降解,随后谷蛋白碱性亚基和球蛋白亚基也逐渐被胃蛋白酶消化降解,而醇溶蛋白亚基则最后被胃蛋白酶作用。随着米糠酸败程度加剧,清蛋白亚基、谷蛋白酸性亚基和球蛋白亚基完全降解的时间先提前后延迟,贮藏3 d米糠制备的米糠蛋白消化产物的亚基条带消失最早,而谷蛋白碱性亚基和醇溶蛋白亚基则总体上表现为更难以被胃蛋白酶消化。通过分子质量分布和粒径分布结果可以看出,米糠酸败过程中米糠蛋白形成了大量共价交联聚集体,且这类氧化聚集体难以被胃蛋白酶消化分解。蛋白质内源荧光峰位结果显示,随着体外胃蛋白酶消化时间延长,米糠蛋白消化产物荧光峰位均发生红移,随着米糠酸败程度增加,米糠蛋白消化产物的红移幅度先增大后减小,在第3天时红移幅度最大。表面疏水性结果显示,随着体外胃蛋白酶消化时间延长,米糠蛋白消化产物的表面疏水性均先上升后下降,在消化20 min左右达到最大值,米糠酸败则会导致米糠蛋白及其消化产物的表面疏水性均下降,这表明米糠酸败对米糠蛋白消化产物中芳香族氨基酸残基和疏水性氨基酸残基的组成、分布及其微环境也产生了重要影响。
[1] 陈正行, 王韧, 王莉, 等. 稻米及其副产品深加工技术研究进展[J].食品与生物技术学报, 2012, 31(4): 355-364. DOI:10.3969/j.issn.1673-1689.2012.04.003.
[2] GUL K, YOUSUF B, SINGH A K, et al. Rice bran: nutritional values and its emerging potential for development of functional food: a review[J]. Bioactive Carbohydrates and Dietary Fibre, 2015, 6(1): 24-30. DOI:10.1016/j.bcdf.2015.06.002.
[3] LOYPIMAI P, MOONGNGARM A, CHOTTANOM P. Impact of stabilization and extraction methods on chemical quality and bioactive compounds of rice bran oil[J]. Emirates Journal of Food and Agriculture, 2015, 27(11): 849-856. DOI:10.9755/ejfa.2015-09-738.
[4] FABIAN C, JU Y H. A review on rice bran protein: its properties and extraction methods[J]. Critical Reviews in Food Science and Nutrition,2011, 51(9): 816-827. DOI:10.1080/10408398.2010.482678.
[5] 吴伟, 蔡勇建, 吴晓娟, 等. 米糠贮藏时间对米糠蛋白结构的影响[J]. 现代食品科技, 2017, 33(1): 173-178; 205. DOI:10.13982/j.mfst.1673-9078.2017.1.027.
[6] 何莉媛, 吴伟, 吴晓娟, 等. 米糠贮藏时间对米糠蛋白体外胃蛋白酶消化性质及其消化产物抗氧化性的影响[J]. 中国油脂, 2017,42(11): 70-74. DOI:10.3969/j.issn.1003-7969.2017.11.017.
[7] RUTHERFURD S M, MONTOYA C A, MOUGHAN P J. Effect of oxidation of dietary proteins with performic acid on true ileal amino acid digestibility as determined in the growing rat[J]. Journal of Agricultural and Food Chemistry, 2014, 62(3): 699-707. DOI:10.1021/jf403146u.
[8] LI Z L, MO L, LE G W, et al. Oxidized casein impairs antioxidant defense system and induces hepatic and renal injury in mice[J].Food and Chemical Toxicology, 2014, 64: 86-93. DOI:10.1016/j.fct.2013.10.039.
[9] ADEBIYI A P, ADEBIYI A O, YAMASHITA J, et al. Purification and characterization of antioxidative peptides derived from rice bran protein hydrolysates[J]. European Food Research and Technology,2009, 228(4): 553-563. DOI:10.1007/s00217-008-0962-3.
[10] ADEBIYI A P, ADEBIYI A O, OGAWA T, et al. Purification and characterisation of antioxidative peptides from unfractionated rice bran protein hydrolysates[J]. International Journal of Food Science and Technology, 2008, 43(1): 35-43. DOI:10.1111/j.1365-2621.2006.01379.x.
[11] BAMDAD F, CHEN L. Antioxidant capacities of fractionated barley hordein hydrolysates in relation to peptide structures[J]. Molecular Nutrition and Food Research, 2013, 57(3): 493-503. DOI:10.1002/mnfr.201200252.
[12] CHEN Nannan, ZHAO Qiangzhong, SUN Weizheng, et al. Effects of malondialdehyde modification on the in vitro digestibility of soy protein isolate[J]. Journal of Agricultural and Food Chemistry, 2013,61(49): 12139-12145. DOI:10.1021/jf404099y.
[13] WU Wei, WU Xiaojuan, HUA Yufei. Structural modification of soy protein by the lipid peroxidation product acrolein[J]. LWTFood Science and Technology, 2010, 43(1): 133-140. DOI:10.1016/j.lwt.2009.05.006.
[14] HUANG Youru, HUA Yufei, QIU Aiyong. Soybean protein aggregation induced by lipoxygenase catalyzed linoleic acid oxidation[J]. Food Research International, 2006, 39(2): 240-249.DOI:10.1016/j.foodres.2005.07.012.
[15] 张敏, 周梅, 王长远. 米糠4 种蛋白质的提取与功能性质[J]. 食品科学, 2013, 34(1): 18-21.
[16] XIA N, WANG J M, GONG Q, et al. Characterization and in vitro digestibility of rice protein prepared by enzyme-assisted microfluidization: comparison to alkaline extraction[J]. Journal of Cereal Science, 2012, 56(2): 482-489. DOI:10.1016/j.jcs.2012.06.008.
[17] 江连洲, 王瑞, 李秋慧, 等. 体外模拟消化过程中大豆蛋白的亚基组成及分子质量分布[J]. 中国食品学报, 2015, 15(10): 65-72.DOI:10.16429/j.1009-7848.2015.10.009.
[18] ZHOU F B, ZHAO M M, CUI C, et al. Influence of linoleic acidinduced oxidative modifications on physicochemical changes and in vitro digestibility of porcine myofibrillar proteins[J]. LWTFood Science and Technology, 2015, 61(2): 414-421. DOI:10.1016/j.lwt.2014.12.037.
[19] SUZUKI Y, ISE K, LI C Y, et al. Volatile components in stored rice[Oryza sativa (L.)] of varieties with and without lipoxygenase-3 in seeds[J]. Journal of Agricultural and Food Chemistry, 1999, 47(3):1119-1124. DOI:10.1021/jf980967a.
[20] YE L, LIAO Y, ZHAO M M, et al. Effect of protein oxidation on the conformational properties of peanut protein isolate[J]. Journal of Chemistry, 2013, 2013: 1-6. DOI:10.1155/2013/423254.
[21] WU W, ZHANG C M, HUA Y F. Structural modification of soy protein by the lipid peroxidation product malondialdehyde[J]. Journal of the Science of Food and Agriculture, 2009, 89(8): 1416-1423.DOI:10.1002/jsfa.3606.
[22] 江连洲, 李佳妮, 邹晓霜, 等. 花青素对大豆蛋白体外胃消化结构的影响[J]. 食品科学, 2018, 39(20): 34-42. DOI:10.7506/spkx1002-6630-201820006.
[23] 刘宇, 董平, 梁兴国. 胃蛋白酶的分离现状及其活性研究进展[J]. 生物学杂志, 2016, 33(3): 75-79. DOI:10.3969/j.issn.2095-1736.2016.03.075.
[24] CHEN Nannan, ZHAO Mouming, SUN Weizheng. Effect of protein oxidation on the in vitro digestibility of soy protein isolate[J]. Food Chemistry, 2013, 141(3): 3224-3229. DOI:10.1016/j.foodchem.2013.05.113.
[25] 尤翔宇, 黄慧敏, 吴晓娟, 等. 过氧自由基氧化对米糠蛋白结构和功能性质的影响[J]. 食品科学, 2019, 40(4): 34-41. DOI:10.7506/spkx1002-6630-20171227-346.
[26] WU W, ZHANG C M, KONG X Z, et al. Oxidative modification of soy protein by peroxyl radicals[J]. Food Chemistry, 2009, 116(1): 295-301. DOI:10.1016/j.foodchem.2009.02.049.
[27] ZHOU F B, ZHAO M M, ZHAO H F, et al. Effects of oxidative modification on gel properties of isolated porcine myofibrillar protein by peroxyl radicals[J]. Meat Science, 2014, 96(4): 1432-1439.DOI:10.1016/j.meatsci.2013.12.001.
[28] CUI Chun, ZHAO Mouming, YUAN Boen, et al. Effect of pH and pepsin limited hydrolysis on the structure and functional properties of soybean protein hydrolysates[J]. Journal of Food Science, 2013,78(12): 1871-1877. DOI:10.1111/1750-3841.12309.
[29] VIVIAN J T, CALLIS P R. Mechanisms of tryptophan fluorescence shifts in proteins[J]. Biophysical Journal, 2001, 80(5): 2093-2109.DOI:10.1016/S0006-3495(01)76183-8.
[30] SIMAT T J, STEINHART H. Oxidation of free tryptophan and tryptophan residues in peptides and proteins[J]. Journal of Agricultural and Food Chemistry, 1998, 46(2): 490-498.
[31] RADHA C, PRAKASH V. Structural and functional properties of heat-processed soybean flour: effect of proteolytic modification[J].Food Science and Technology International, 2009, 15(5): 453-463.DOI:10.1177/1082013209350347.
[32] BAMDAD F, WU J P, CHEN L Y. Effects of enzymatic hydrolysis on molecular structure and antioxidant activity of barley hordein[J].Journal of Cereal Science, 2011, 54(1): 20-28. DOI:10.1016/j.jcs.2011.01.006.
Effect of Rice Bran Rancidity on Structural Characteristics of in Vitro Pepsin Digest of Rice Bran Protein
WU Wei, HE Liyuan, HUANG Huimin, et al. Effect of rice bran rancidity on structural characteristics of in vitro pepsin digest of rice bran protein[J]. Food Science, 2019, 40(17): 14-21. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-20180925-262. http://www.spkx.net.cn